[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]
Chapter 7
CRANIAL NERVES
FINAL COMMON PATHWAY
Key Words: oculomotor, trochlear, abducens, mesencephalon, pons, medulla oblongata, tonic, burst, burst-tonic
Outline
VI. Cranial Nerves - final common pathway controlling eye movements
- Introduction
- Cranial Nerves III, IV & VI
Ipsilateral and contralateral projections & location in brainstem
- Motoneuron Response
Only respond to position information- velocity has been integrated previously
- Characteristics of Motoneurons
Classifications: Tonic; Burst; Burst-Tonic
On-Off directions- firing rate increases in on-field
Eye position dependent thresholds
Recruitment
Introduction
Cranial nerves form final common pathway. There is no specialization of neuron type and type of EM. e.g. same cells in III activate MR during saccades, pursuits convergence , VOR and OKN. (Classes of eye movements are specialized prior to the cranial nerves in premotor nuclei.) The following two figures are presented as a review of cranial nerves III, IV, and VI, and their associated nuclei:
Fig 7.1, Table 7.1, |
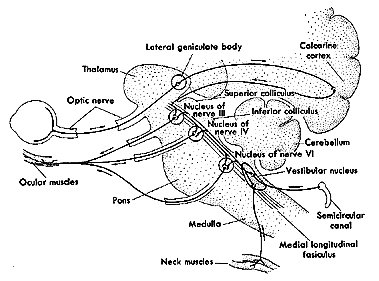 |
Medulla Oblangata |
Pons |
Mesencephalon (midbrain) |
VIII (vestibular) |
VI (abducens) |
III (oculomotor), IV (trochlear) |

Cranial Nerves III, IV, & VI and Nuclei
There are twelve pairs of cranial nerves altogether. They are numbered in rostral to caudal sequence. The oculomotor (III), trochlear (IV), and abducens (VI) nerves innervate the six extraocular muscles, iris and ciliary body. The acronym LR6SO4 may help you memorize the name of a nucleus and its associated number. The abducens (VI) nerve innervates the ipsilateral lateral rectus. The trochlear (IV) nerve innervates the contralateral superior oblique. The oculomotor (III) nerve innervates the rest.
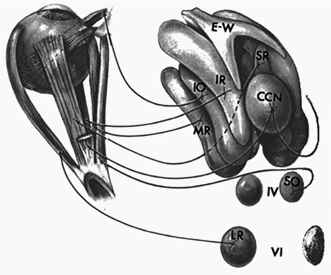 |
Fig 7.3 |
Clearly, The Oculomotor nucleus (III) is the most complex of the cranial nuclei associated with eye movements. From this nucleus the medial rectus, inferior rectus, and inferior oblique are innervated ipsilaterally, whereas the superior rectus is innervated contralaterally. The anterior portion of the oculomotor nucleus also contains innervation for pupil and accommodation in a specialized region called the Edinger Westphal nucleus. Afferents from this nucleus synapse in the ciliary ganglion prior to innervating their target muscles. The regions of the oculomotor nucleus that control position of various eye muscles are illustrated in figure 5-48 on page 137 of Adler. (see pages 135-143 in Adler).
Fig 7.4 DN: Dorsal Nucleus |
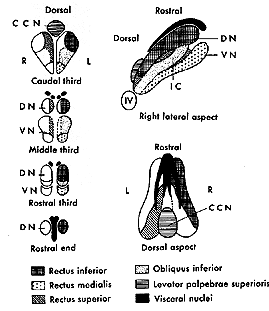 |
Cranial nerves III, IV, and VI represent the final common pathway for all classes of eye movements. All cells whose axons project from these nerves carry information for saccades, pursuits, vergence, VOR and OKN.
The tables below list the crossed and uncrossed innervations of the 3 motor nuclei and the paired yoked muscles in the two eyes. Note that pairs of yoked muscles are innervated by the same side of the brainstem. i.e. the SO and IR are innervated from one half of the brainstem and the IO and SR are innervated by the same half of the brainstem. The LR and MR are both innervated by the abducens nucleus via an interneuron that originates from the abducens and projects to III on the contralateral side.
Table 7.2 Projections of Motor Nuclei
Ipsilateral |
Contralateral |
|
III Oculomotor |
MR, IO, IR |
SR |
IV Trochlear |
SO |
|
VI Abducens |
LR |
MR via interneuron |
Table 7.3 Yoked Muscle Pairs
Direction of Movement |
Extraocular Muscle (OS) |
Extraocular Muscle (OD) |
LEFT |
LR |
MR |
RIGHT |
MR |
LR |
UP (in left gaze) |
SR |
IO |
DOWN (in left gaze) |
IR |
SO |
UP (in right gaze) |
IO |
SR |
DOWN (in right gaze) |
SO |
IR |
Motoneuron Response
Cells within each of the nuclei that project axons along the cranial nerves are called motoneurons. They are the final common pathway for all classes of eye movements, and their main function is to control the position of the eye, that is they encode position information that can change in time to result in changes in eye position. This is true for pursuits and saccades where position signals are varied over time. Position signals are derived by supranuclear regions that transform velocity to position in regions called neural integrators.
Fig 7.5 Neural integration of position information from velocity information takes place in supranuclear regions (See Chapter 9.) |
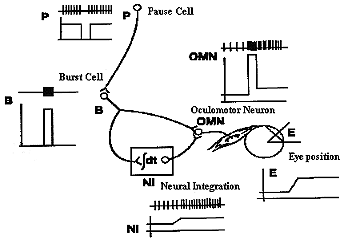 |
Characteristics of Motor Neurons
Muscles increase their contractile force either by increasing firing rate of individual motoneurons or by recruitment of more motoneurons to stimulate the muscle. Motoneurons can be divided into three functional categories. These are tonic, burst and burst-tonic units. The latter two categories are also referred to as phasic. Tonic cells are thought to be responsible for holding the eyes steady or moving them slowly, and the more phasic or burst like motoneurons are responsible for moving the eyes rapidly. Tonic-burst have both tonic and phasic properties. All motoneurons have the following characteristics (reference: Adler p. 141 and Leigh and Zee pp. 180-181):
- They have clear on-off directions (they increase their firing rate in the direction of agonist activity).
- All cells participate in all classes of eye movements including steady fixation, even in the primary position of gaze.
- Each cell (especially tonic) has an eye position threshold at which it begins to fire. Cells with low thresholds begin firing when the cell is in the off field of the muscle. Cells with high thresholds begin to fire after the eye has moved past the primary position by as much as 10 degrees into the on field of the muscle. A group of tonic cells for the medial rectus have thresholds that range from low to high. This range is the basis for recruitment of cells when it is necessary to increase the contractile force of a muscle. For example, when the eye moves into a muscle's field of action, there is an increase in the number of cell axons innervating the muscle. The graded threshold of motoneurons accomplishes this recruitment of active cells as the eyes move into the on position or field of action for the muscle.
- The other way in which contraction is increased is by increasing the frequency of spike potentials for a given neuron. Once their threshold is exceeded all cells increase their firing rate as the eye moves further along in the on direction of the muscle. Different cells increase their firing rate linearly as the eye moves into the on field.
Firing rate can be plotted as a function of eye position. (See Fig 7.6 below) The slope of these functions describes the cell's sensitivity. Cells whose firing rate increase slowly with eye position have low sensitivities to eye position whereas cells that increase their firing rate abruptly have high sensitivities.
Fig 7.6 Note illustration B, which plots firing rate as a function of eye position. The slope describes the motorneuronís sensitivity. |
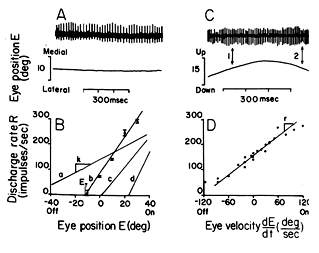 |
A general observation for most motoneurons (especially tonic) is that sensitivity and threshold tend to be correlated. That is, cells with low thresholds have low sensitivities. They don't change their firing rates dramatically as the eye moves into their field of action. Cells with high thresholds tend to have high sensitivities. This means that once their threshold is exceeded, their firing rate increases abruptly with change in eye position. The net effect of this is that contraction increases mainly as a result of recruitment when starting from the off position of a muscle, and it increases mainly as a result of increased firing rates of individual motoneurons when it moves beyond the primary position of gaze into the on field of the muscle.
The summed activity of all motoneurons as the eye moves from its off field to the upper limit of its on field increases exponentially as a result of recruitment. This exponential increase in necessary because different amounts of force are needed to make small changes in eye position starting from different orbital positions. It takes little change in innervation to move the eye away from its off position because of the elastic force of the stretched agonist and slackened position of the antagonist. In contrast it takes a greater change in innervation to move the eye the same amount when starting from a position near the full limits of its potential contraction. This is due in part to the spring force of the stretched antagonist and the loss of stretch of the agonist. Thus the innervational pattern compensates for the mechanical properties of the agonist-antagonist pair of muscles they innervate.
Review Questions:
- Why are the cranial nerves called the final common pathway?
- What are the functions of tonic, burst and tonic-burst cells?
- How do threshold and sensitivity characterize the activity of tonic moto-neurons?
[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]