[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]
Chapter 6
MUSCLE MORPHOLOGY AND MECHANICS
Key Words: Myofibril structure, sliding filament theory, controllers with spindles and stretch receptors, mechanical models (acceleration, velocity, position, mass, viscosity, elasticity)
Outline
V. Anatomy, Morphology, and Mechanics of EOM (contí)
- Introduction
- Morphology of 5 types of Muscle Fibers
Striated muscle fibers, myofibrils, sarcoplasmic reticulum, t-tubules
Sarcomeres, myosin (thick) and actin (thin) filaments, I and A bands, Z & M lines
Actin- tropomyosin, G & F actin, torponin
- Muscle Contraction
Sliding filament theory
Neurochemical events during contraction
Functional types of EOM fibers
- Muscle Force Mechanics - 3 factors
Inertia - load (mass) antagonist and globe- limits acceleration, force acceleration curve- inertia constant (J)
Viscosity - limits velocity, force velocity curve- viscosity constant (B)
Spring elasticity (series-tendon, and parallel-muscle sheath)- limits length, length tension curve- spring stiffness constant (K). K increases with contraction and changes point of equilibrium
- Muscle Receptors
Spindles - stretch receptors sense length - parallel, intrafusal
Golgi tendon organs - sense force - serial attachment
Introduction
In order to understand the innervational patterns that are needed to shape different types of eye movements, it is helpful to be familiar with the structure and function of ocular muscle fibers. The mechanical properties of muscle produce a viscosity that limits the speed of contraction and requires specific strategies by the CNS for controlling fast eye movements. Much of this material is typically presented in an ocular anatomy or physiology class. This is an intentional review of that material.
Morphology of Muscle Fibers
Gross structure
Extraocular muscle is a specialized form of striated muscle. It is called striated because of the dark and light bands or striations running perpendicular to the long axis of the muscle. The large muscle is composed of fibers which are in turn composed of cylindrical bundles of myofibrils. Each myofibril has a diameter of 1 micron. Each myofibril is enveloped in a self contained bag called endoplasmic reticulum. This envelope is interwoven with another self-contained network of channels called the T-tubule system. The T-tubules convey action potentials throughout the interior of the myofibril to insure simultaneous contraction of all regions of the muscle.
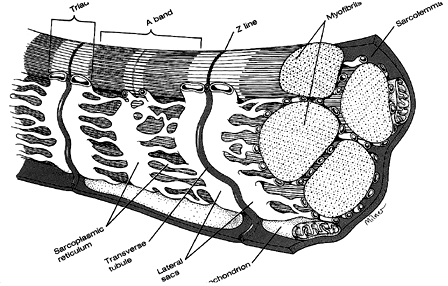 |
Fig 6.1 Structure of striated muscle. |
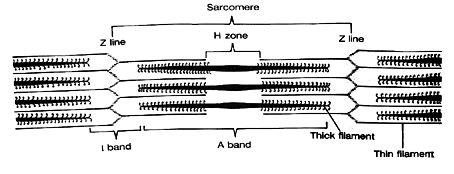
Myofibrils are composed of regions of light and dark bands. The light bands (called I bands) are isometric to polarized light (meaning they transmit polarized light). The darker bands (called A bands) are anisometropic to polarized light (meaning they do not transmit polarized light). A given section of striation is called the sarcomere, which is the basic contractile unit of striated muscle.
The density of striations results from the interdigitation of two types of parallel filaments. Thin filaments (called actin) lie in the I band. Thick filaments (called myosin) lie in the A band. These two types of filaments are interdigitated such that during contraction they slide over one another, which causes a shortening of the sarcomere. Because the sarcomeres are serially arranged, the overall contraction of the myofibril equals the summed activity of individual sarcomeres.
The A band is divided into several zones of darkness that result from varying degrees of interdigitization of actin and myosin. In the center of the A band is a lighter region called the H zone containing only myosin. Adjacent to this, the actin filaments partially overlap the myosin filaments and cause further darkening. The light center of the A band is where the actin has yet to slide during contraction. The very light I bands occur in regions containing only actin. The sarcomere is demarcated by dark lines (called Z lines) which are anchor points for actin being pulled in opposite directions toward the center of the sarcomere. The center of the A band has an M line which represents a thickened region of the myosin fiber.
Fig 6.2 Detail of muscle fiber |
 |
|
 |
Substructure of thin and thick filaments.
The thin filaments are made up of three proteins. 1) The actin has a core made up of a long strand of protein called tropomyosin. 2) These strands are surrounded by a coil of globular spherical sub units called G-actin. G-actin forms a double chain like a twisted pearl necklace called F-actin. F stands for fibrous and G for globular. These serve as binding sites for cross bridges between actin and myosin filaments. 3) Small globs called troponin are attached to the tropomyosin. These act to block the attachment of myosin with actin when contraction is not occurring.
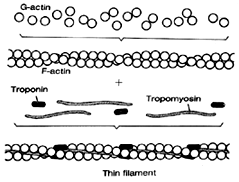 |
Fig 6.4 Protein structure of a thin filament. |
The thick myosin filament is like a golf bag. It is made up of about 250 protein strands that look like golf clubs. (See Fig 6.3) All the arms of the clubs are bound into a common shaft and the club heads point out perpendicular to the shaft. The heads emerge in a helix that spirals around the shaft. When the myosin overlaps with actin filaments, each myosin shaft is surrounded by a hexagon of 6 actin filaments.

Fig 6.5 Overview of muscle structure.
The Sliding Filament theory
In 1954 Andrew Huxley proposed that the myosin heads attached with the G actin pearls on the actin filament forming a cross bridge and then the myosin heads underwent a shape or conformational change that exerted a pulling force on the actin filament causing it to slide over the myosin filament. At the end of a brief twitch contraction the head detached, the bridge was broken and the process began over again with another twitch. Many successive twitches, each changing length by 4 nanometers, caused an overall contraction of the muscle.
Neural chemical events during contraction
Contraction begins with an action potential along the motor axon that reaches the neuromuscular or motor end plate. This causes the release of acetylcholine and depolarization of the muscle fiber. The depolarization is propagated throughout the myofibril by the t-tubular system and causes a change in permeability of the sarcoplasmic reticulum. Calcium is then released into the interstitial spaces of the muscle. Calcium acts as a clutch that determines the linkage between myosin heads and actin filaments. Four changes occur with the release of calcium:
- Troponin has a high affinity for calcium and it pulls back the troponin to expose G-actin sites where myosin heads can attach and form cross bridges. This constitutes a structural or conformal change of the actin filament.
- The contraction is fueled by ATP which is produced in mitochondrion by the breakdown of glucose. ATP combines with the myosin head and forms a charged intermediate that attaches with an actin subunit on the thin filament after calcium unveils the g-actin sites. This is the formation of the cross bridge.
- The third step is the release of energy by splitting ATP into ADP and an inorganic phosphate. The release of energy causes another conformal or shape change in which the myosin head rotates like a ratchet gear and slides toward the center of the sarcomere and causes actin to slide forward about 4 nanometers.
- The twitch contraction ends when calcium is withdrawn by a calcium pump back into the sarcoplasmic reticulum. Then the troponin gates shut by covering the g-actin sites and the cross bridges are broken momentarily.
Another twitch sequence begins and contraction consists of a rapid series of twitches that can be several hundred per second lasting throughout the change in muscle length. Unlike most skeletal muscle, the EOM are always contracting. They never relax. Thus the EOM are always undergoing these electro-chemical and bio-mechanical actions throughout our life.
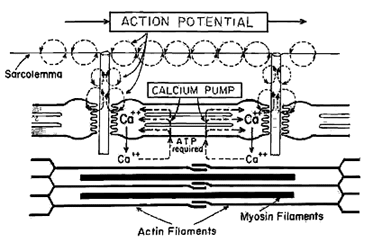 |
Fig 6.6 Schematic representation of events which initiate muscle contraction. An action potential at a motor end-plate releases acetylecholine. Resulting depolarizing propagates throught the t-tubule system, causing calcium to be released by the sarcoplasmic reticulum. |
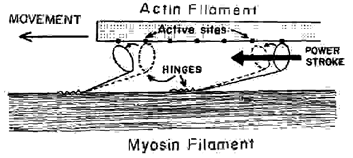 |
Fig 6.7 Conformational change in which the myosin head rotates and causes the actin filament to slide about 4nm. |
Contractile force
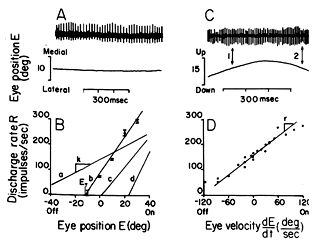
There are two ways to increase the strength of contraction. One is firing frequency and the other is recruitment. Regarding firing rate, a single action potential that triggers a twitch contraction lasts only 1-3 msec. However a single twitch contraction of the muscle resulting from this brief innervation lasts 10-100 msec due to the slow uptake of calcium after the formation of cross bridges. Therefore successive action potentials may activate the muscle fiber before it relaxes and summate with the previous twitch. The amount of force generated by the muscle increases with firing frequency until the responses fuse in what is called fused tetanus. Ripple from the unfused firing rates doesn't produce a rippled response because individual muscle fibers are averaged with asynchronous responses of other fibers. In addition the stretch receptors help smooth or dampen the response as we will see later.
Fig 6.8 Releationship between discharge rate of motor axon and position/ velocity of eye. (The amount of force generated by a muscle increases with firing frequency.) |
 |
The second means of increasing contraction force is to increase the number of motor axons that stimulate the muscle via recruitment. Initially contraction begins with stimulation by only the neurons that respond tonically for long durations but not too strongly. As the demand for increased contraction goes up, more neurons stimulate the muscle with even higher frequency bursts. These neurons tend to be phasic and fatigue easily. Weíll see physiological records of this recruitment phenomenon when we go over the motoneurons of the third cranial nerve.
Three types of extraocular muscle fibers
Three functional types of fiber have been identified in extraocular muscle. All three are active at all times and are recruited differentially during different ocular movements. The first class is singly innervated twitch fibers (SI). These are specialized for rapid contraction and respond to increased frequencies of spike potentials up to 175-300 Hz. They are most suited for initiating rapid saccadic eye movements. They also play a role in fixation and pursuit conditions. The second class of fiber is the multiply innervated non-conducting fibers (MINC) which are specialized for tonic longstanding contraction. They do not conduct action potentials because the Ca++ is released from surface membrane rather than the interior tubular system and sarcoplasmic reticulum. They respond to low frequency spike potentials of 50 Hz. The third class of fiber is multiply innervated fibers with action potentials (MIC). These fibers respond to lower stimulus spike frequencies than do SI fibers and they conduct at slower velocities. They are rare in comparison to other fiber types. The multi innervated fibers are responsible for the majority of tonic activity in all gaze positions including the primary position.
Muscle Force Mechanics
The contractile force depends on the degree of overlap of myosin and actin fibers. Initially there is increasing overlap of thick and thin filaments and muscle force increases with continued contraction. Force reaches a peak when the actin strands originating from opposite Z bands meet. The contractile force decreases with further sliding as opposite actin filaments begin to overlap. Additional factors go into this length tension curve.
Muscle also exhibits spring like properties that determine the magnitude of contraction or position of the eye in the orbit. The cross bridges can flex, and tendons and connective tissue have elastic properties. Also the muscle becomes stiffer as contraction proceeds. A spring is a mechanical device that responds to increase in length by generating a restoring force. This force is proportional to length, as shown in the following equation:
F = stiffness (K) * length (L)
where K is the spring or stiffness constant. A stiff spring or muscle has a greater restoring force than a soft spring.
The force needed by an agonist will depend on the length of the antagonist. The more a muscle is stretched the greater the restoring force. As the antagonist is stretched, as when eye position is in the field of action of the agonist, its restoring force increases. At the same time its stiffness is decreased slightly with decreased contraction. The position of the eye is determined by an equilibrium point between spring forces of the agonist and antagonist. The length-tension curve applies to contraction. Adler's text book shows a different length tension curve that represents the passive restoring force for a detached extraocular muscle. The purpose of this curve is to measure muscle stiffness. Surgeons measure this to determine how much to recess a muscle to correct strabismus.
These interactions are complicated when describing movement rather than position because of the lengthening of an antagonist muscle during contraction of an agonist. This requires about twice as much force compared to isometric contractions. This is because cross bridges are being broken during the lengthening to stretch the muscle and the stiffness of cross bridges is quite high. Accordingly, the innervation the muscle must be adjusted differently during static eye position and while making a movement past the same eye position. This property is measured as a physical analog called viscosity. If we had no viscosity we would only need to generate a force to begin rotation of the eye, much like spinning a marble, and then we would need a breaking force to stop the spin. But the muscles do have viscosity and viscosity requires an increase of force for an increase velocity. The more viscous a medium the more force needed to move through it at a given velocity, as indicated in the following equation:
F= viscosity (B) * velocity
Think of two different viscous liquids, soup and oatmeal porridge. If you want to stir them with a spoon, it takes a lot more force to stir the oatmeal than the soup. The extraocular muscles have high viscosity coefficients so that a great deal of force is needed to make quick movements such as during saccades. In order to make these fast movements that may approach 1000 degrees/second, extraordinary forces are needed compared to the forces required during pursuits of much lower velocity.
A third mechanical factor that limits motion is inertia or mass, as shown in the equation:
F = inertia (J)* acceleration (A).
This determines how rapidly the eye can change its velocity or accelerate. Since the eye has little mass this is a very small factor in the overall force program that regulates eye movements.
The two main factors that are viscosity (B) and elasticity (K). These determine velocity and position of the eye respectively. The total force depends upon all three physical factors, mass, viscosity and stiffness.
F= (J * A )+ (B * V) + (K * X)
Muscle receptors
Normal skeletal muscle behavior requires coordinated activity between several groups of muscles. There needs to be coordinated actions of agonist and antagonists and between yoked muscles of the two eyes. In addition there needs to be precise damping of eye muscles at the end of very abrupt and rapid movements such as saccades. These actions are assisted by feedback generated with sensors within the muscle. There are two types of receptors in EOM that I will describe. One is the stretch receptor called the muscle spindle. These are attached in parallel with the muscle along its belly and they monitor changes in muscle length and velocity. The other sensor is the Golgi tendon organ that is attached in series at the tendon of the muscle. These monitor muscle force and prevent excessive force from tearing muscle by breaking innervation. Both of these receptors are highly prevalent in human extraocular muscle. They are less prevalent in monkey and again are prevalent in goat. Their role in eye movements is still not well understood. We will review their general role in skeletal muscle and then look at their possible role in eye movements.
Muscle spindles
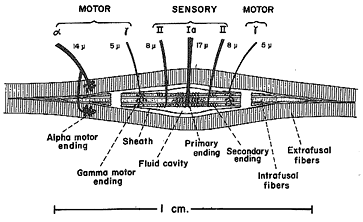 |
Fig 6.9 |
Muscle spindles are composed of several striated muscle fibers that are separate from the main body of the muscle. These fibers are called intrafusal whereas the main muscle fibers are called extrafusal. Each spindle is built of 3 - 10 intrafusal muscle fibers that are attached at their ends to the main extrafusal muscle. These intrafusal fibers contract just like extrafusal fibers. Intrafusals are innervated by gamma efferent fusimotor fibers whereas extrafusal fibers are innervated by the alpha efferent skeletal motor system. Normally the two types of fibers are stimulated equally and the spindle lengthens and shortens with the rest of the muscle. However if an external load causes a passive lengthening of the extrafusal fibers, this stretches the intrafusal fibers without a corresponding decrease in innervation. This passive stretch is sensed by the central region of the spindle that has sensory or afferent fibers that wrap around the spindle and whose firing rate increases when the spindle is lengthened. These afferent signals pass to the brainstem and elsewhere in the brain and can result in a reflex contraction of the extrafusal muscle via the alpha efferent system to minimize the lengthening. This automated response is called the load or stretch reflex.
Basically, this reflex maintains the status quo of the muscle. This reflex is also called the damping reflex. It can be demonstrated by striking the knee to stretch the patellar tendon of the quadriceps muscle. The result of this passive lengthening is a sudden contraction of the quadriceps and a knee jerk reflex. Normally we don't need a stretch reflex in EOM because there are no unexpected loads placed on eye like those that occur with other skeletal muscle. The only time the eyes might have any dramatic changes in load is during blinks when the eyelids move it back into the orbit by as much as a millimeter. During this blink all of the extraocular muscles co-contract, possibly in response to the stretching of the muscles.
Proprioceptive information from human extraocular muscle passes to the brain via the ophthalmic branch of the 5th or trigeminal nerve. These impulses are projected throughout the brain to sites including the brainstem, cerebellum, superior colliculus, LGN, and visual cortex and they may be used for a variety of purposes. Laboratory attempts to demonstrate a stretch reflex of the EOM during saccades have not been successful. This is probably because a stretch reflex would interfere with a saccade and it is normally suppressed during the saccade. High velocity saccades must overcome the high viscosity of the extraocular muscles. To do this the muscle is hit with a very abrupt and strong increase in innervation called the burst. This can only be achieved if the stretch reflex is inhibited. Without the stretch receptors the reciprocal inhibition depends upon the organization of the brain stem connections. The classic stretch reflex may be used when the eye stops at a new position and the force needs to be braked. Viscosity helps braking along with a very low eye mass, reverberations do occur, much like the oscillations that make a bell ring after you strike it with a hammer. The eye oscillations would be an interaction between eye mass and muscle elasticity. The muscle spindles could dampen these mechanical oscillation because they are not accompanied by alpha efferent oscillations and the dynamic stretch reflex would smooth them out. Because the eyes move in response to a rapid sequence of twitches, we also need the mechanical and physiological dampening functions to ensure smooth continuous movements during pursuits. It is possible that stretch reflex is utilized by the cerebellum to modify the late trajectory of saccades.
The stretch reflex is also known to assist the coordinated actions of agonists and antagonists. When a muscle spindle is stretched, there is both increased innervation to the agonist and decreased innervation to the antagonist. This interaction reduces the load from the agonist imposed by the antagonist. This reciprocal inhibition is called Descartes-Sherrington's law of reciprocal innervation. Descartes invented the afferent-efferent reflex and came up with the reciprocal innervation law in 1626. Sherrington tested the law in 1890. It may not always require a stretch reflex because there are interneurons in the brainstem that coordinate stimulation of one muscle with inhibition of its antagonist. For the oculomotor system Sherrington' Law of reciprocal inhibition refers to interactions between the agonist and antagonist within the same eye.
Control of eye position requires stretch receptors in the absence of light. Gross nystagmus is observed in darkness when stretch receptor afferents are blocked by cutting the 5th nerve in a cat. Stretch receptors also appear necessary to maintain precise conjugacy of the two eyes movements. When there is a herpes infection of the facial nerve (tic-delareux), the symptoms are relieved by cutting the 5th nerve. This de-innervates the afferent path for proprioceptive impulses from extraocular muscle. Interestingly when this is done we lose the fine control of binocular conjugacy. Finally, cats can't visually guide paw placement with their eyes if the 5th nerve has been cut.
Golgi Tendon Organs
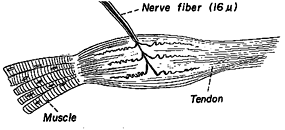 |
Fig 6.10 |
Golgi tendon organs are force detectors located in series with the muscle in its tendon. This receptor receives no innervation but sends an afferent signal into the brainstem when force is applied to the muscle against the tendon. The resulting tendon reflex or lengthening reaction is to inhibit the alpha innervation to the muscle applying the force. The purpose of this reflex is to prevent a muscle from being torn. Personal experiences involve what are called "stingers" in which a limb goes limp when a tendon is struck, or when a muscle gives way when it is overloaded. Again, this reflex should be suppressed during saccades when the eye needs abrupt changes in force to overcome the high viscosity of the eye.
Review Questions:
1. Describe the architecture of striated muscle.
2. Describe the sliding filament theory.
3. What are the roles of calcium, troponin, and ATP in muscle contraction?
4. What factors contribute to the acceleration, velocity and length of muscle?
5. Explain the difference between muscle spindles and Golgi tendons.
[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]