[Table of Contents] [Home] [Glossary] [Video] [Help]
CROSS-COUPLING
OF ACCOMODATION AND CONVERGENCE (AC/A and CA/C)
Key words: accommodation, convergence, adaptation,
gradient, calculated, tonic, phasic
Outline
XII. Near Response: Pupil constriction, Accommodation
and Vergence.
Part III: Cross-coupling (of accommodation and convergence)
- Introduction (Examples of cross-coupling)
cyclotorsion and convergence, AC/A, CA/C, cross-axis VOR, VOR gain & vergence, compensation for noncomitance
- Measurement of AC/A Ratio
Gradient- stimulus and response measures
Calculated
- Linearity
- Stability
age affects
drug affects
Adpatation
- Dynamics
Phasic stimulation
interactions with tonic adaptation
- Innervation of convergence and accommodation
Introduction
Frequently many motor responses occur simultaneously
when we perform complex tasks such as seeing. We need to
aim the eyes in the direction of the target to permit foveation,
converge correctly to align targets on corresponding retinal
points, control the focus to sharpen the foveal image, control
pupil size to adjust luminance, and compensate for head and body
movements during fixation. All of these tasks are performed
simultaneously whenever we change fixation from one object to
another. To help coordinate these motor responses, the brain
stem has many associated reflexes such that stimulation of just
one of these motor activities causes the others to respond appropriately
as though they had been stimulated directly. Generally these
cross-coupled responses serve to make coarse adjustments in associated
gaze shifts and then retinotopic cues are used to refine the
response.
The most well known of the visual cross-coupling responses is the near triad whereby stimulation of either convergence or accommodation causes changes in convergence, accommodation and pupil size. This association is called a synkinesis. This is not the only synkinesis in the visual system. Also the VOR gain is related to the state of convergence. As you translate your head laterally, the amount your eyes need to counter rotate to maintain exact fixation increases as viewing distance decreases. The VOR automatically increases with convergence. Another cross-coupling response occurs between cyclovergence and eye elevation. When we look down at the ground plane our eyes automatically excycloverge and when we look up they automatically incycloverge. This helps us fuse the ground and ceiling planes which are declinated and inclinated with respect to the visual axes in these vertical positions. Another example is the counter roll of the eyes when the head rolls to one side, stimulating the otoliths. Clearly there are many other examples outside of vision that involve coordination of heart contraction, digestion, and postural control.
In Optometry, we are primarily concerned with the cross-coupling between accommodation and convergence. When accommodation is stimulated alone by occluding one eye, the covered eye still converges via a coupling referred to as the AC/A ratio. Similarly, if we converge with pinhole pupils we stimulate vergence but accommodation also responds. This cross-link is referred to as the CA/C ratio. These ratios are normally tuned to be very accurate and appropriate for the geometry of our eye separation, but occasionally something goes wrong and the ratios are either too high or too low. This causes imbalance of convergence and accommodation such that we may over-converge when we accommodate because of a high AC/A ratio. This induces an esophoria. If we are also hyperopic, it may induce an esotropia. When this occurs one of the main treatments is to prescribe bifocals to reduce the accommodative stimulus and the resulting esophoria. However many surgeons, particularly in Europe, like to do surgery on these patients. The result is that they create exotropes at far after they diverge the eyes because the eyes are too divergent when accommodation is not stimulated. It is very important to understand the source of the esotropia in a child to avoid making this mistake. Now letís look at the normal status of the AC/A.
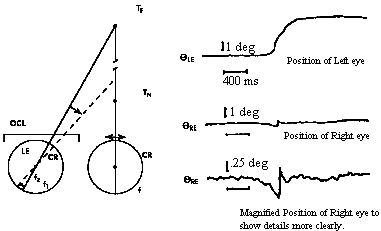
|
Fig 20.1 |
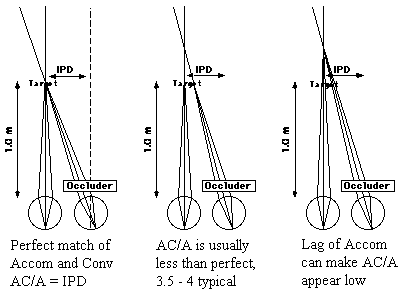
|
Fig 20.2 |
Measuring Amplitude of the AC/A
The AC/A ratio is measured by two techniques called
the calculated AC/A and the gradient AC/A. The
gradient AC/A is measured by occluding one eye, placing a minus
one lenses over the open eye and measuring the change in phoria
as shown by deviation of the covered eye. This can be done
by alternate cover test or a diplopia test such as red lens or
Maddox rod. The resulting change in convergence stimulated
by the change of accommodation quantifies the gradient AC/A ratio.
The calculated AC/A ratio is measured by comparing the near
and far phorias using the equation:
calculated AC/A ratio =IPD + (near phoria - far phoria)/MA
IPD is the interpupillary distance measured in centimeters. Phoria
is measured in prism diopters. MA (meter angles) = 1/near
target distance measured in meters. Ideally, the phoria
is zero at all distances. In the ideal case, the calculated
AC/A = IPD in centimeters. For example if you converge to
1 meter, your eyes form an angle of convergence of 6 prism diopters
(assuming an IPD of 60mm) or whatever your IPD is in prism diopters. If
you are more eso at near than far, the ideal AC/A is increased
by the additional change in phoria between the far and near test
distances divided by the change in accommodative stimulus from
far to near. Generally, the calculated AC/A is slightly
higher than the gradient because of the addition of proximal
convergence to the measurement of the phoria.
The gradient AC/A can be specified as the ratio of the change
in phoria divided by the accommodative stimulus, in which case
it is called the stimulus gradient AC/A ratio, or by the
ratio of the change in phoria divided by the accommodative response,
in which case it is called the response AC/A ratio. The
normal value of the stimulus ratio is 3.6 prism diopters/D accommodation
and the normal response AC/A ratio is 4.0 prism diopters/D. The
difference between stimulus and response AC/A is due to the lazy
lag of accommodation which reduces the denominator of the response
ratio.
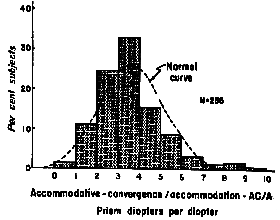
|
Fig 20.3 |
Linearity
If you measure the gradient AC/A ratio with different amounts of accommodative stimulus (e.g. 1 or 2 or 3D) or at different distances corresponding to different starting levels of accommodation, there is little change of the AC/A ratio because it is linear as long as you stay in the linear branch of the accommodation stimulus-response function. (Linearity means that if the accommodation stimulus is doubled, the resulting amount of convergence should also double.)
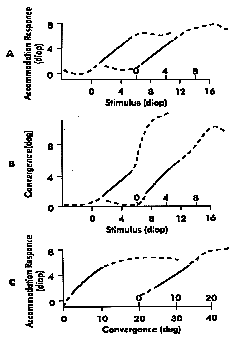
|
Fig 20.4 A) Accommodative response as function of stimulus Notice that plot C indicates that the AC/A ratio is linear, at least in regions where accommodation response is linear. |
Stability of AC/A
Age effects
The AC/A ratio remains very stable throughout life. Its day to day measurements usually vary by less than 0.25 prism diopters. This is rather amazing since the IPD changes radically from birth to about 6 years of age when there is rapid growth of the cranium. Yet the AC/A doesn't change.
|
Fig 20.5 |
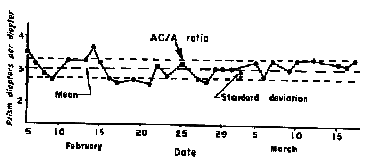
|
Drugs
Drugs affecting the autonomic nervous system also cause temporary changes in the AC/A. Parasympatholytic drugs such as atropine or cyclogyl that block the parasympathetic branch of the autonomic nervous system by inhibiting uptake of acetylcholine via competition cause a marked reduction of accommodative amplitude and corresponding increases in the response gradient AC/A ratio. The AC/A ratio increased from 3.5 to over 20/1 in only 2.5 hours after the instillation of homatropine and it takes over 20 hours for it to return to normal. In contrast, the clinical applications of milder and shorter acting parasympatholytic drugs such as Mydriacyl, Tropicamide, and Cyclogyl (cyclopentolate) which cause a reduction of accommodation and increase in the AC/A ratio in only 40 minutes and wears off in about 2 hours. You can also decrease the stimulus AC/A with the prescription of Miotics which increase the depth of focus by decreasing pupil size. This has been used to treat accommodative esotropia in patients such as very young children who will not comply with wearing glasses.
|
Fig 20.6 (AC/A ratio indicated with bold line. Near point of accommodation shown with dashed line.) |
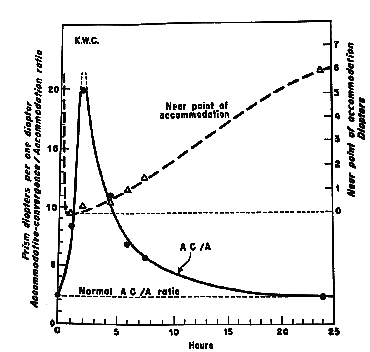
|
Dynamics of the AC/A ratio
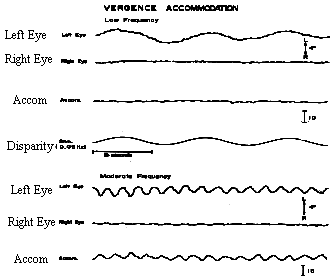
What happens when we change accommodation continuously with sinusoidal or pendular variations in dioptric power? Is the amount of AC/A the same as the gradient measure which stimulates accommodation with various step changes in lens power? One way to test this is to monocularly track a sinusoidal variation of the stimulus to accommodation and measure the dynamic changes in convergence of the covered eye. When this is done, the AC/A varies dramatically with the temporal frequency of the sinusoidal stimulus. When accommodation changes very slowly, such as 1 diopter in 10 seconds, the AC/A can be as low as zero. When accommodation changes more rapidly, such as 1 D in 1 second, the AC/A shows its normal measured amplitude. This velocity dependence on the value of the AC/A ratio suggests that at slow velocities, tonic accommodation is adapting to the accommodative stimulus and tonic accommodation does not have a cross-link with accommodative vergence. However at higher velocities the accommodative response is mainly phasic and there is a large amount of AC/A. These results suggest that phasic retinal-blur-driven reflex accommodation stimulates accommodation vergence but adaptable tonic accommodation does not. This can be summarized with a block diagram or schematic of the cross-coupling between accommodation and vergence.
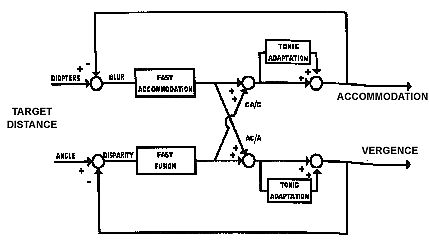
|
Fig 20.7 Cross-links of accommodation and convergence |
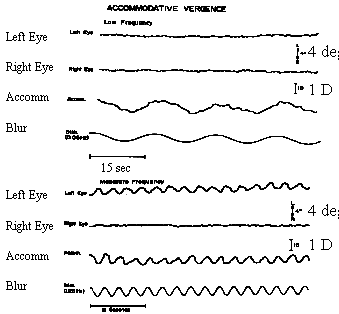
|
Fig 20.8 Note that for very low frequencies, the AC/A is zero, since tonic accommodation is not cross-linked. However, for higher frequencies, the accommodative response is mainly phasic, and the AC/A ratio is normal.
|
|
Fig 20.9 |
If both CA/C ratio and AC/A ratios are velocity sensitive, you may wonder what keeps the AC and CA active during steady fixation when there are no changes in stimuli to accommodation or convergence. Focus and eye alignment are maintained by an adaptation response of both systems. The accommodative system adapts to replace CA and convergence adapts to replace AC. This is shown in the cross-link block diagram. An imbalance of adaptablilty of accommodation and convergence can lead to abnormally large and small cross-link ratios that interfere with normal binocular vision.
Abnormal values of CA/C ratio and AC/A ratios tend to be reciprocally related. A person with a high AC/A usually has a low CA/C. This reciprocal relationship is due to a single causal factor which is an imbalance of tonic adaptation of accommodation and convergence. For example, assume that accommodation is highly adaptable but convergence is not. This means that CA will build up a tonic aftereffect of accommodation but AC will not build up a tonic aftereffect of convergence. In addition, due to the lack of adaptation of convergence, the phasic component of vergence must always be active and this causes more than normal stimulation of CA. The result is a very high CA/C ratio and very low AC/A ratio.
Similarly, if vergence is very adaptable and accommodation is not, the lack of accommodation adaptation causes phasic accommodation to be overly active and cause excess innervation of AC. This is exacerbated by the robust adaptation of convergence that enhances the already large AC. The highly adaptable convergence will lower phasic vergence and reduce the CA to below-normal values. One treatment is to perform eye exercises to enhance the weakened adapter so that it matches the strength of the stronger adapter and balances the interactions between accommodation and convergence.
Innervation of Convergence
Both convergence and accommodation are stimulated by the oculomotor
nucleus. This nucleus receives pre-motor inputs from a variety
of supranuclear regions of the brainstem and cerebellum. It used
to be thought there was a convergence center called the nucleus
of Perlia near the Edigner Westfall nucleus but this has not
been verified. Instead there is a main source of innervation
from a region dorsal to the III in the mesencephalon called the
supraoculomotor nucleus. This nucleus contains cells for convergence
and accommodation that are homologous to the pause, burst and
tonic cells described earlier for saccades. Burst cells encode
vergence velocity and tonic cells code vergence amplitude or
position. There are both convergence and divergence burst neurons
but convergent cells are more abundant. The tonic cells have
firing rates that vary with vergence amplitude. The firing rate
is related only to vergence response and not to right or left
eye version position, nor to vertical eye position. The response
of a given cell is plotted as a function of vergence angle. It
shows that each cell has a slope or sensitivity and threshold.
The thresholds are all within a few degrees of orthophoria and
the slopes are all very similar. Presumably the tonic cells represent
the neural integration of the burst cells. Some of these tonic
cells exhibit prism adaptation aftereffects. They continue to
respond when the disparity stimulus had been removed by covering
one eye and at the same time there is a convergence aftereffect.
Some cells respond to disparity, others to accommodation and
some to both. These cells represent the final common pathway
for all vergence responses to disparity, accommodation, and probably
proximity and tonicity.
Two other regions have also been found with cells that are responsive
during accommodation and vergence. On is in the posterior interposed
nucleus of the cerebellum (one of the deep cerebellar nuclei)
and the other is in the NRTP, nucleus reticularis tegminti pontis.
Not much else is known about them.
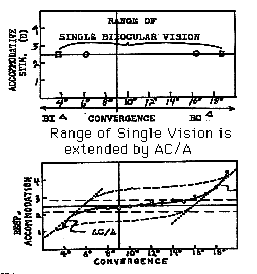
Clinical Application of Maddox Components
Chapter 22 describes how the various Maddox components are measured
and plotted on a single graph to illustrate their additivity.
The graph is used to determine how a given patient utilizes the
various components of convergence to obtain clear single binocular
vision. The figure below is a graph showing a plot of convergence
on the horizontal axis and accommodation on the vertical axis.
The circles represent the limit of clear single vision and the
squares represent an extension of the range of single vision
by exerting accommodative vergence. The result is a greater range
of singleness, but with blurred vision.
|
|
Review Questions:
1.How are the gradient and calculated AC/A ratios computed and how do they differ in amplitude ?
2.What are the stimulus and response AC/A ratios.
3.How does the CA/C ratio change with age?
4.What is the effect of a parasympatholytic drug on the AC/A ratio?[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]