[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]
Chapter 19
VERGENCE EYE MOVEMENTS
Key words: Maddox, tonic, proximal, disparity, accommodative
Outline
XII. Near Response: Pupil constriction, Accommodation and Vergence.
Part II: Convergence
- Introduction
- Units of Measurement- prism diopters, degrees, meter angles
- Maddox components
Tonic
Proximal (Coarse adjustment)
Disparity (Fine adjustment)
Accommodative (See chapter 21)
- Vergence Dynamics
Asymmetric vergence
Symmetrical convergence
- Effects of prisms and lenses
- Neural Control of Vergence
Introduction
Disjunctive or vergence eye movements are movements of the eyes in opposite directions. They can be horizontal, vertical and cyclo. These serve the same function as conjugate movements of the eyes which is to preserve the alignment of the two retinal images on corresponding retinal points. Vergence is required when changing viewing distance, whereas versions accomplish the task when changing direction of gaze. Vergence doesn't appear in animals with laterally placed eyes but these animals do exhibit conjugate movements because the VOR stabilizes both eyes no matter where they point in space.
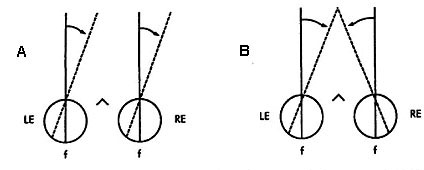 |
Fig 19.1 |
 |
Fig 19.2 |
Units of Measurement
Vergence is measured in degrees and prism diopters. The meter angle is also used in order to get a comparable metric for accommodation and convergence. Meter angles equal the reciprocal of the viewing distance and the value is approximately equal to the accommodative stimulus in diopters.
MA = 1/(target distance in meters)
Meter angles (MA) can be converted into prism diopters simply by multiplying it by the interpupillary distance (IPD) in cm:
Prism Diopters = IPD (cm) * MA
Maddox Classification of Vergence
At the turn of the century, Maddox described vergence as a linear summation of responses to a variety of stimuli. In other words, he suggested there were separate response components of convergence to these stimuli which summed linearly to form the composite response. As we will see, all components of vergence are not active at the same time. Some components are coarse adjustments and are responsible for setting up a baseline or beginning point for the other fine adjustment components which refine binocular eye alignment. Some components are specialized for shifting attention abruptly, some for maintaining fixation on a given target, and some are responsible for coordination with other near response motor functions. The vergence components described by Maddox that serve these functions are 1) tonic, 2) proximal, 3) fusional and 4) accommodative. The first three are discussed in this chapter. Accommodative vergence is discussed in Chapter 20.
Maddox Classification of Vergence |
|
Tonic vergence
When we are born and during the first 6 weeks of life, our eyes diverge during sleep. The eyes start out exotropic and then become aligned. When we die the eyes also diverge. This divergence posture is known as the anatomical position of rest and it represents the vergence angle in the absence of any innervation. As we develop during the first 1.5 months of life, the resting vergence angle is reduced to nearly zero by the development of tonic vergence which reduces the resting position from the anatomical position of rest to the physiological position of rest. This is the angle that vergence assumes when we are awake and alert, in the absence of any stimulus to accommodation or binocular fusion or perceived distance. Usually, this physiological position of rest differs from the vergence demand, (i.e., the amount of vergence needed to fuse a distance target). The discrepancy between the vergence demand at a far viewing distance and the physiological position of rest is called the phoria. The phoria is an error of binocular alignment, but the error is only manifest during monocular viewing or any condition that disrupts binocular vision. If binocular vision is allowed (not disrupted) the other components of vergence reduce the error nearly to zero.
Tonic or baseline vergence is adaptable. It can be modified to compensate for overall changes in binocular alignment caused by accommodative convergence, prism in spectacles, and misalignment caused by injury and normal developmental changes such as widening of the interpupillary distance (IPD). These changes are responsible for the nearly constant phoria that is present throughout life. The phoria distribution in the general population is Kertotic (Tait) with a small exo mean. (See Fig 19.3) The maintenance of a near zero phoria is called orthophorization which is analogous to the emmetropization process for refractive error. As stated above, tonic vergence develops postnatally. Children have exotropia in sleep until they are 6 weeks old. Similar changes are seen in cats. It appears there is an automatic mechanism that converges the eyes in infants until fusion signals there is binocular alignment. If fusion is not signaled then the eyes continue to converge into esotropia. For example, if you put a vertical prism over one eye or occlude one eye of a newborn kitten, the eyes keep converging past parallel and the cat becomes esotropic. However if the cat is allowed binocular vision, when the eyes reach alignment they stop converging. The tonic mechanism appears to be part of a normal developmental sequence in which convergence is increased and stops when binocular cells signal alignment.
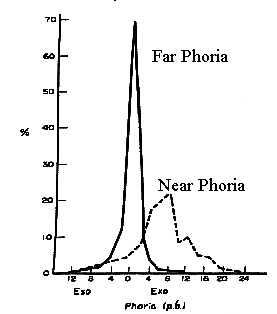 |
Fig 19.3 |
This adaptive process continues throughout life. We need to compensate for the growth of the IPD. The wider our eyes become, the more we need to converge for near targets. There are also problems of noncomitance. Muscle innervation can be traumatized and disrupt the pattern of innervation that results in yoking described by Hering's Law. When a paresis first appears it is localized. It may gradually disappear or become more uniform in all directions of gaze. This phenomenon is referred to as spread of comitance. Anisometropic spectacle corrections also disrupt the normal alignment pattern needed for fusion. For example a person wearing a plus lens over the right eye needs to over converge at near and over diverge at far. Our oculomotor system is able to make these non-comitant adjustments such that the phoria varies with direction and distance of gaze consistent with the problems encountered.
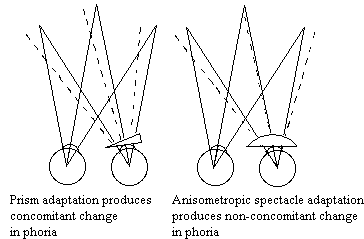 |
Fig 19.4 Solid lines show orthophoric alignment before adapatation. Dashed lines show how the right eye will be deviated after adaptation, the left eye viewing. |
There are two components of the adaptation process that compensate for eye misalignment. One is typically called prism adaptation and it refers to our ability to adjust our phoria in response to a uniform prism produced by displacement of the optical center of a lens away from the visual axis. This prism is constant throughout the visual field and it produces a global or constant change in phoria in all directions of gaze. The global adaptation is very rapid and can be complete in several minutes in a normal patient. Patients who are unable to perform this adaptation will benefit from the prescription of prism to reduce their phorias.
The second component of adaptation is a local process that adjusts the phoria separately for each direction and distance of gaze. For example, a patient with an "A" pattern is more esophoric or tropic in up than down gaze. Successful local adaptation will reduce the eso in up gaze without introducing exophoria in down gaze. This spread of comitance takes longer than the global adaptation described above. It can occur in 1-2 hours or in months for large non-comitant deviations. Local adaptation can be facilitated by partial field prism prescriptions that reduce the phoria and allow adaptation to respond to a smaller demand. Once the eyes have adapted, the prism can be reduced until adaptation controls the entire oculomotor deviation.
Finally, the phoria may vary with viewing distance because of an unfortunate combination of excessive accommodation associated with hyperopia and a high AC/A ratio. This combination can cause esophoria at near.
The tonic vergence adapter can compensate for the extreme demands on the sensory-motor fusion process by reducing the phoria at the problematic viewing distance. It relieves the stress on fusional vergence by reducing the phoria.
Adaptation appears to be in response to the continued activity of fusional vergence. Initially, we use fusional vergence to overcome the phoria error. If we fuse for several minutes, tonic vergence adapts and relieves fusional vergence. However on the surface it appears as though nothing is happening because the eyes are constantly aligned throughout this adaptation process. Internally, the control of vergence alignment is shifted from a fast acting or phasic component to a slow acting tonic level of control. The tonic component adapts the vergence to such stimuli as prism.
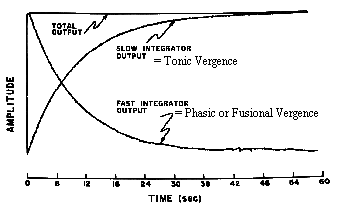 |
Fig 19.5 Initially, fusional vergence corrects for the error. Tonic vergence then gradually adapts and relieves fusional vergence. |
In the absence of adaptation ability, prism is prescribed. Optometrists prescribe a prism to relieve the inability to comfortably fuse. A problem that sometimes occurs is that the patient adapts to the prism and exhibits the same phoria through the prism as they had before it was prescribed. It is as though the system adapted back its original phoria. This adaptation phenomenon is called prism adaptation. This topic will be considered again below under the heading of fixation disparity.
Methods for Measuring the Phoria |
|
Proximal Vergence
The term proximal refers to perceived distance. Perceived distance is classified as a spatiotopic cue because it results in a conscious percept of distance and direction. Our perception of space is made up of an infinite number of different targets in different directions and distances. Our eyes can only select and fuse one of these target configurations at a time. One of the main tasks of the oculomotor system is to select one fixation target from many alternatives. This selection process usually results in a shift of binocular fixation from one distance and direction to another. Proximal vergence serves as a course adjustment mechanism and accounts for nearly 100% of the eye movement when gaze shifts are too large for us to sense them from blur and disparity. We perform these large gaze shifts without needing to see the new intended target clearly or even with both eyes. We only need to have an estimate of where that target is using monocular cues for distance and direction. The majority of binocular shifts in fixation are stimulated by proximal vergence. Once the shift has been made another component of vergence (fusional vergence) takes over to maintain the response at the new target location.
Disparity vergence
Fusional or disparity vergence is analogous to optical reflex accommodation. It serves as a fine adjustment mechanism that holds or maintains fixation of a given target and corrects for minor changes of disparity caused by small movements of the head or the target being inspected. Disparity vergence is the most studied component of vergence and a disproportional amount of emphasis has been placed on it over the other components. It is important, but it is not the main vergence mechanism. (Proximal vergence is the dominant vergence mechanism.) The greatest feature of disparity vergence is that it utilizes feedback in the form of retinal image disparity that is needed to align the eyes accurately and to instruct the adaptation mechanism of any corrections needed in tonic vergence.
Vergence dynamics
Most studies of vergence dynamics involve stimulating pure convergence or symmetrical vergence along the midline, however in real life most vergence is combined with lateral shifts of gaze. We shift gaze diagonally in space following a pattern of asymmetric vergence where one eye needs to move more than the other. The dynamics of symmetrical and asymmetric vergence are different from one another and we will discuss them separately.
Asymmetric vergence (Interactions between versions and vergence)
Asymmetric vergence is a combination of lateral saccades with horizontal vergence. The velocity of vergence is combined with saccades to yield some very high velocity vergence responses that appear to result from yoked saccades of unequal amplitude. The vergence velocity resulting from differences in the two saccades can approach 50 deg/sec. It is not clear if these movements are the result of a combination of symmetrical vergence with equal saccades, or if we are able to make saccades of unequal amplitude. It is likely that both of these explanations are correct.
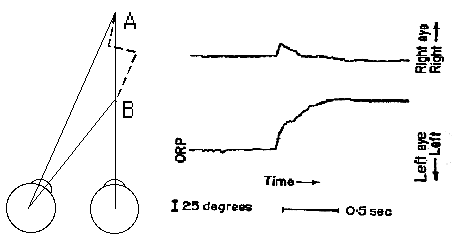 |
Fig 19.6 |
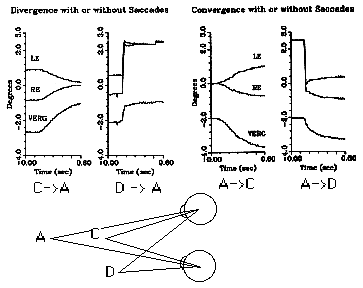 |
Fig 19.7 Notice that asymmetric vergence (A to D, or D to A) is faster than asymmetric vergence. |
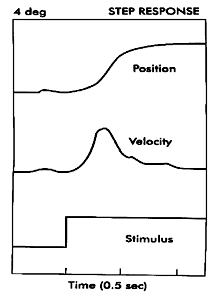
Symmetrical Convergence
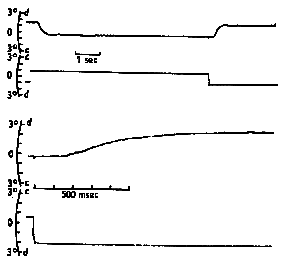
Symmetrical vergence movements tend to be more sluggish than asymmetric vergence. They have latencies of 160 msec which is shorter than a saccade and about the same as a pursuit. Their response time is sluggish taking .8 to over 1 second for completion. When the vergence is stimulated abruptly by a prism, the step response resembles a decaying exponential. (See Figs. 19.8 and 19.9 below) Its initial velocity is proportional to the magnitude of the disparity and then as the response proceeds it slows down smoothly and comes to rest at the new desired vergence angle. Rashbass and Westheimer showed the relationship between initial velocity and stimulus amplitude. Velocity increases with amplitude up to a limit of 1-2 degrees. There is a limited range of pure small disparities to which we can respond without the help of other distance cues. If the disparity exceeds 1-2 degrees it is out of the range for disparity processing and other distance cues are required stimulate vergence.
 |
 |
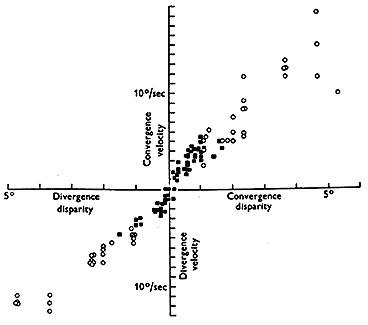 |
Fig 19.10 |
The disparity vergence system is used continuously to track smooth or slow variations of the disparity stimulus caused for example by body sway and posture instability. Our ability to make these movements is tested by having subjects track sinusoidal variations in target disparity. We are fairly good at this for tracking frequencies up to 0.5 - 1.0 Hz and then the response amplitude falls off. We can track higher frequencies only if the amplitude of the stimulus is very small. This is a velocity limit of several degrees per second.
The disparity vergence system has some prediction capability. Without prediction we would expect the vergence response to lag behind the stimulus by 1 latency period, but it keeps up the stimulus as long as its motion is predictable. Prediction is even seen in the prism step response. Without it, you would expect the eyes to overshoot the target and continue responding for one latency period. You would also expect unsteady oscillations of attempted secondary corrections of the overshoot. However, the eyes are able to slow down as they approach their final destination and stop on target at the end of the step response because we anticipate when our eyes will reach their goal. This anticipatory function is performed by the cerebellum for most body movements. Prediction also helps slow down the step response and make it a decaying exponential. During the response the stimulus disparity is gradually being reduced. The shrinking disparity causes the subsequent velocity of vergence to be reduced. Finally, the disparity is zero and the response should terminate. Prediction is necessary to get the response to terminate without waiting an additional latency period. As mentioned before prediction helps dampen possible unsteady responses that would occur because of the latency period.
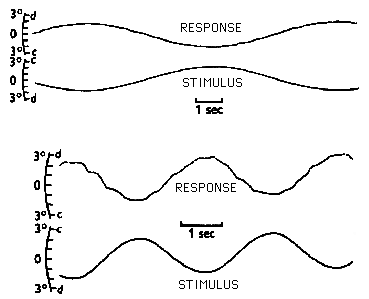 |
Fig 19.11 |
Effects of Prisms and Lenses on vergence and phoria
Both prisms and lenses produce optical image displacements. The amount of displacement produced by a lens can be calculated from Prentice's rule (Prism diopters = lens eccentricity in cm * diopters). The vergence posture induced by a prism or lens determines the direction of the phoria. For example, fusional or disparity convergence is stimulated by a base-out prism and results in exo-phoria. However, accommodative convergence stimulated by a minus lens causes eso-phoria. Similarly, fusional divergence stimulated by a base-in prism causes eso-phoria, and accommodative divergence stimulated by a plus lens causes exo-phoria.
Neural Control of Vergence
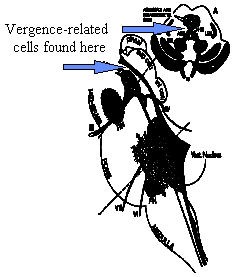
The neural control of vergence is only understood at the final common pathway in the brainstem. Here the supraoculomotor nucleus region that contain near response cells controls afference from accommodation and disparity and tonic inputs as they are relayed to the third nerve nuclues. The supraoculomotor nucleus contains burst and tonic cells as well as phasic-tonic cells that are characteristic of the saccadic pathways. It is thought that velocity signals related to disparity stimlui innervate the bursters and that velocity information is integrated to form the position signal that is processed by the tonic cells. Accommodative-vergence is already represented by these cells so that the wiring for cross-coupling between accommodation and convergence must occur more centrally in the pathways.
Fig 19.12 Regions involved in vergence are indicated by arrows. |
 |
Current anatomical studies are discovering other regions of the brainstem, and parts of the cerebellum that are involved in the control of convergence and MRI studies will likely localize cortical sites involved in vergence control.
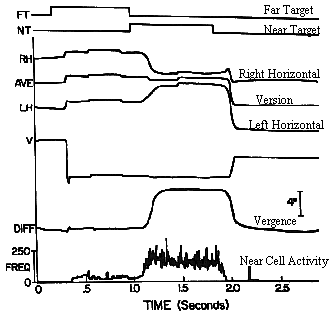 |
Fig 19.13 (Fixation first at a far target, and then at a near target). |
Review Questions:
- What is the transformation from prism diopters to meter angles.
- List four general methods for dissociating binocular vision to conduct phoria tests.
- What are coarse and fine adjustment components of vergence?
- List the 4 Maddox components of vergence
- What are differences in the dynamics of symmetrical and asymmetrical vergence?
[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]