[Previous Chapter] [Next Chapter]
[Table of Contents] [Home] [Glossary] [Video] [Help]
[Previous Chapter] [Next Chapter] |
Chapter 14 Key words: saccades, latency, main sequence, pulse, step, integration
Thus we need to select the stimulus to which we wish to respond, ignore or suppress responses to compelling gaze stabilization stimuli such as head motion and optic flow, and at the same time remain prepared to shift the motor response to unexpected urgent stimuli such as large looming objects. All of this requires selective voluntary attention, the ability of fixation and tracking responses to override stabilization reflexes, and low level reflexes to call our attention to novel stimuli. All of these activities are carried out at the same time by various high level centers in the brain. As a result there appear to be multiple pathways controlling the same type of eye movement. Some of these pathways are concerned with attention and voluntary control of eye movements such as reading and in visual search, and some with reflexive attention shifts to novel stimuli, and some with interactions between tracking and stabilization and fixation eye movements such as the fast phase of OKN and the VOR. |
|
|
||||||||||||||||
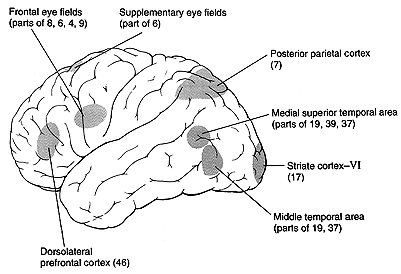
The exact purpose of many of the pathways that control eye movement is still not completely understood. In addition there are species differences, so that the pathways are different in cats, rabbits, monkeys, and human. This chapter will present the pathways that are most understood from the occipital and frontal eye fields and the superior colliculus, but Leigh and Zee point out anatomical studies of many other regions of the brain that appear to be part of the oculomotor pathways. This chapter reviews the function and pathways for tracking eye movements. The functions of tracking eye movements, which include visual grasping and gaze holding, are accomplished by saccades and pursuits respectively. These classes of eye movements are distinguished by their function, speed, control characteristics, and anatomical sites for their regulation. |
|
|||||||||||||||||
|
|||||||||||||||||
|
|||||||||||||||||
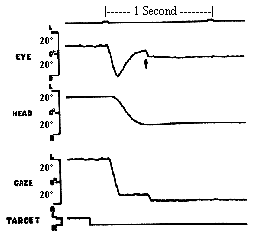
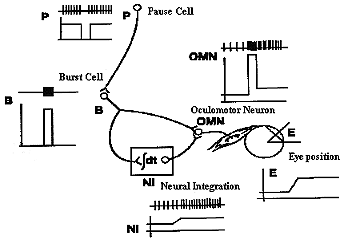
Saccades get the eye to a direction in the orbit as quickly as possible with little regard for the conservation of energy. It's a survival tactic. Consider a baseball player who is stealing second. He starts off with an incredible effort to speed up to the second base and then breaks as fast as possible to avoid being tagged out. The effort is much greater than if he had simply walked or jogged over to second. The saccadic system does the same thing. It accelerates as quickly as possible to overcome the small mass and large viscosity of the eye and orbital structures, and then adopts a new level of innervation to maintain a new position which is restrained by elastic forces of the ocular muscles. The rapid acceleration is accomplished by the frequency of the pulse, the amplitude by the duration of the pulse, and the new sustained eye position by the amplitude of the step innervation programmed by the PPRF. In addition these actions require reciprocal inhibition of antagonists during the movement. While the agonist constricts, the antagonist is totally inhibited, and at the end of the movement the antagonist becomes active to help break the saccade when the antagonist innervation is reactivated to its new step level corresponding to the new eye position. |
|||||||||||||||||
|
|||||||||||||||||
Most of this innervational pattern is preprogrammed before the saccade actually occurs, much like a ballistic missile is preprogrammed. After the launch, the missile follows its course determined by preprogrammed forces. However, interactions with unforeseen forces, like shearing winds, can occur. This is unlike the guided missiles that take in feedback during their flight to make corrections for unforeseen events or for a path which was too complex to be programmed prior to the launch. Saccades have been referred to as ballistic because their force program of pulse and step appeared to be preprogrammed one latency period prior to the response. |
|||||||||||||||||
|
|||||||||||||||||
|
|||||||||||||||||
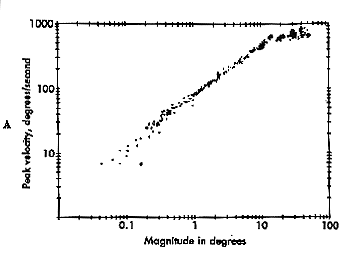
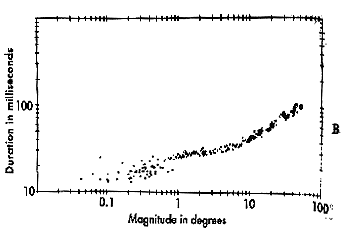
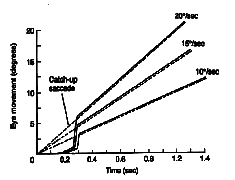
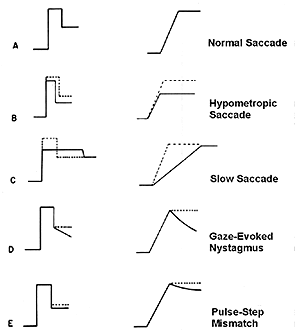
Pulse-Step mismatches These observations are summed up in the main sequence diagram which is a continuous function or envelope that describes the duration and velocity of normal saccades based upon their amplitude. |
|||||||||||||||||
|
Fig 14.8 and 14.9 The main sequence. |
||||||||||||||||||||
In a main sequence diagram, saccadic duration and velocity are plotted as a function of saccadic amplitude. The main sequence diagram was adopted by Bahill and Stark from the Hertzsprung-Russell diagram of star classification, which plots luminosity versus temperature. Most stars lie along the diagonal of this plot. Likewise, most saccades lie diagonally on the duration versus amplitude and velocity versus amplitude plots. The main sequence diagram reflects the pulse component of the saccade, and it can be used as a diagnostic tool to evaluate the bursters of the midbrain. If saccades are sluggish, their main sequence values will be low for the amplitude of the resulting saccade. |
||||||||||||||||||||
|
||||||||||||||||||||
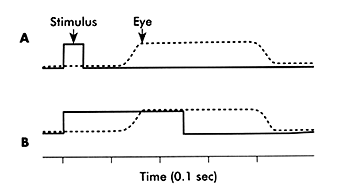
Westheimer showed that if you presented a target that jumps from the fovea to a new retinal location and promptly jumps back to it in less than 100msec, the eye responds with the expected latency of 200 msec after both displacements had occurred. The saccade moved the eye away from the target on the fovea. Then after 200 msec the eye looked back to the target. The interval between saccades was independent of the interval between the target jumps away and back to the fovea. This suggested that the saccadic system could only react to one target at a time and there was a fixed latency or refractory period, during which a second saccade could not be initiated after the first. Finally it suggested that no new information could be taken in during the refractory period. More recently however there have been experiments demonstrating that the eye does pay attention to the target during the first 110 msec of the latency period so that if multiple displacements in the target do occur they can affect the amplitude of the initial saccade response, and even cause additional short latency supplementary saccades to follow up in 20 msec rather than 200 msec. Some studies have shown that you can even change the direction of a saccade during flight if a second stimulus is presented late in the refractory period. Therefore the saccade is not really ballistic because it takes in information during its latency period. A likely alternative explination is that two separate saccades were programmed in parallel and there times of execution overlapped one another. It is possible that whole volleys of saccades are programmed in advance of moving to a new target site. There is an initial hypometric saccade that has a normal 200 msec latency and subsequently there are corrective saccades with latencies as short as 20 msec. This could be the nature of express saccades. This prediction ability is likely to be a function performed by the cerebellum since it normally predicts the completion of most motor movements in order to coordinate walking running and speech. Similarly, the cerebellum may be involved in some way with terminating the saccade at the right moment by sensing any errors in amplitude and activating the antagonist as a break in the case of potential overshoots. To do this we need to predict when the saccade will end. If we don't make this prediction, we end up with saccade dysmetria and over- and under-shoots. |
||||||||||||||||||||
|
||||||||||||||||||||
Saccadic Suppression and Spatial direction constancy Corollary discharge signals are likely to come from the neural integrators of the brain stem. This computation occurs in the parietal lobe and takes some time, maybe as long as 50 msec. To avoid transient errors in perceived direction, the visual percept is suppressed momentarily during the saccade. You can experience this by looking back and forth between the reflected images of your eyes in a mirror. You never see your eyes move. The suppression begins about 50 msec before you begin the movement and ends about 30 msec after the movement. The suppression is not total. It is due to three mechanisms. The first mechanism is the mechanical optical smearing of the retinal image during the saccade. The second process is a 20% reduction of visual sensitivity, and the third and strongest effect is visual backward masking in which the perception of a prior target is suppressed by a later one. Its as though the stronger stimulus at the end of the saccade overtakes the prior weaker stimulus during the saccade and suppresses it within the visual pathways. The same suppression effect is seen without a saccade simply by jerking or shifting the retinal image. Problems in this suppression and the general integration of eye position and retinal image position result in apparent motion of the world during eye movements. This condition is called oscillopsia and it can affect patients with recently acquired nystagmus. However, after several weeks or months the oscillopsia usually disappears when the eye-brain learns to anticipate the rapid nystagmoid movements of the eyes. The oscillopsia probably results from a mismatch in sensed retinal image motion and eye motion sensed via efference copy. When the perceptual system recalibrates to match retinal and extra-retinal motion, oscillopsia disappears. |
||||||||||||||||||||
|
||||||||||||||||||||
|
||||||||||||||||||||
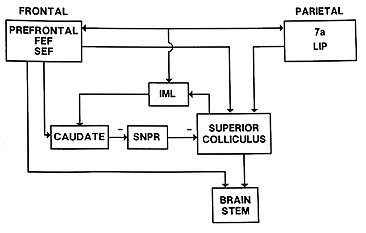
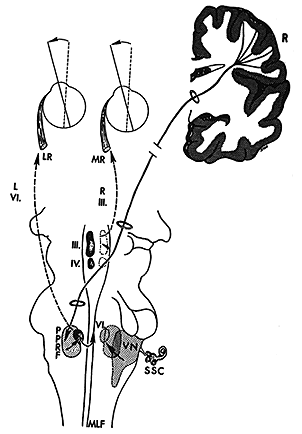
The FEF project two main efferent pathways for the control of saccades, one to the superior colliculus and the other to the PPRF and riMLF. The fibers from the frontal eye fields descend from the cortical field to ipsilateral superior colliculus and, cross the midline, to the contralateral PPRF. Gaze palsies or lesions of the PPRF cause drift of the eyes to the contralateral side and an inability to look to the same side. The FEF specifies to the SC the amplitude and direction of an upcoming saccade. It also participates in the process of maintaining and releasing fixation. Neither the colliculus or FEF are exclusively required to generate saccades. Either one can be ablated and saccades continue. However if both are ablated, saccades are abolished. Thus both regions receive some independent afference and the FEF has several pathways to the brainstem to generate saccades. Lesions of the FEF may prevent patients from making voluntary saccades to remembered target locations without a strong visual stimulus to guide them. This condition is known as pseudo-ophthalmoplegia. |
||||||||||||||||||||
|
||||||||||||||||||||
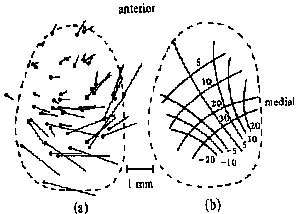
Superior colliculus control of saccades The colliculus has several layers. The superficial layer contains a visual sensory map of the visual field that is derived from both direct retinal projections via the optic tract and cortical projections. The intermediate and deep layers contains a motor map of the visual field. It contains cells with movement fields that are much more organized than those of the FEF. These cells respond to all sensory modalities including vision, audition, and tactile. The direction or spatial location of all of these sensory stimuli are mapped in the colliculus relative to the fovea. Like the FEF, stimulation of one side of the SC causes a conjugate saccade to the contralateral side and stimulation of both sides is necessary to evoke purely vertical saccades. The motor map has a polar organization and is relative to the fovea so that it signals change of fixation from the current eye position and not to a particular orbital location. The larger the retinal error to be corrected the more caudal is the point of activity. Thus small saccades are stimulated rostrally and large saccades are stimulated caudally with the accompaniment of head movements. Medial stimulation causes upward components and temporal stimulation causes downward components of saccades. Pure vertical only occurs with bilateral stimulation. A current theory being tested is that during a saccade, as the retinal error is reduced, the activity in the colliculus travels rostrally toward the fixation pole as a traveling wave. The output of the SC projects to the longlead bursters of the PPRF and the riMLF for horizontal and vertical components of saccades respectively. |
||||||||||||||||||||
|
||||||||||||||||||||
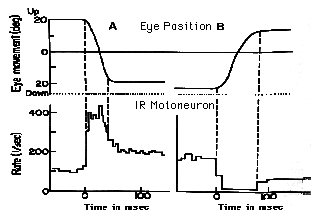
Superior Colliculus: Fixation |
||||||||||||||||||||
|
||||||||||||||||||||
Review Questions:
|
||||||||||||||||||||
[Previous Chapter] [Next Chapter] |
||||||||||||||||||||