[Previous Chapter] [Next Chapter] |
Chapter 13 Key words: VOR, OKR, otoliths, saccule, utricle
In its current state the vestibular system detects position and motion of the head in space by integrating information from peripheral receptors located in the inner ear on either side of the head. This provides a sense of equilibrium that is not conscious but is considered as the sixth sense. The sense is essential for the coordination of motor responses, eye movements, and posture. Without it, we would be unable to move about without total confusion concerning posture or ability to see objects clearly. Anomalies of this organ lead to inability to stand or walk. There can be nausea, vomiting, abnormal eye movements (nystagmus) and sensory-neural hearing loss due to imbalances in the endolymph. These symptoms make up a condition called Meniere's syndrome. |
|
The vestibulo-ocular reflex is very sensitive and can respond to angular accelerations or decelerations as small as 0.5 deg/sec. This reflex is made up of 3 synapses consisting of the vestibular bipolar cells of the vestibular nerve near the mechanical transducers, the vestibular neurons of the brainstem, and the cranial nerves controlling innervation to the extraocular muscles. The speed of this reflex is easily demonstrated by comparing the visibility of your finger as you attempt to track it as you move it back and forth at 4 Hz compared to holding the finger steady and shaking your head at the same rate. The finger is much more visible with the head shake than the pursuit tracking response. Pursuits have an upper frequency response of 1-2 Hz and the VOR can respond at least twice that frequency. Recall that the extraocular muscles primarily respond to position signals but the vestibular stimulus is acceleration. In order to obtain the eye-position-related signal found in oculomotor neurons, a twofold integration (acceleration to velocity to position) has to take place. One integration is determined mechanically by the capula-endolymph system. The endolymph movement translates acceleration into velocity. Accordingly a head velocity signal comes to the vestibular nerve. The second integration (to a position signal) takes place centrally in the neural integrator of the vestibular nuclei. Structure of the mechanical transducer Organization of the Cupula, Hair Cells |
|
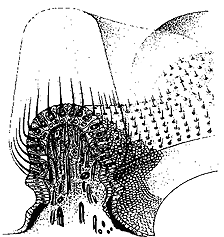
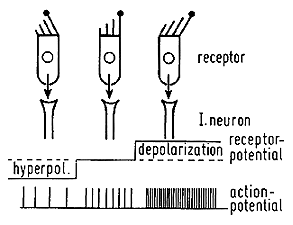
When the head is rotated, the inertia of the fluid in the semicircular duct causes the fluid to push against the cupula, producing an angular displacement of the sensory hairs of the receptor cells. This distortion either causes either a depolarization and stimulation of the vestibular nerve or hyper polarization and inhibition of the vestibular nerve. Due to the organization of hair cells, the horizontal canals are stimulated when the endolymph is moved toward the ampulla and the hair cells in the vertical canals are depolarized when the endolymph moves away from the ampulla. This means the right horizontal canal causes increased vestibular activity when the head rotates to the right, and the anterior canal causes increased activity with downward head rotation, and the posterior canal causes increased activity when the head is rotated upward. |
|
||||

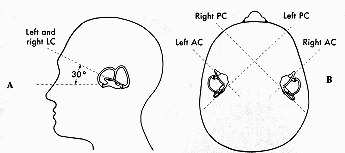
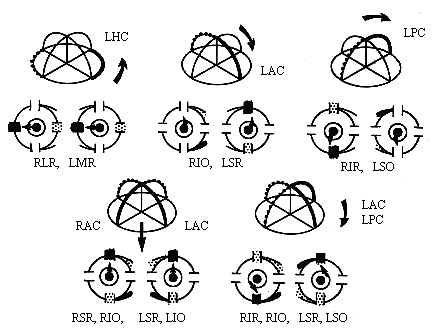
There are three canals that are paired on the two halves of the head. The horizontal canals are paired and the anterior canal on one side is paired with the meridian of the posterior canal on the contralateral side. (See Fig 13.6) These are opponent pairs so that when one canal is stimulated by a given head rotation its paired member on the contralateral side is inhibited. For example, downward head motion causes increased firing of the vestibular nerve for the anterior canal on one side and decreased firing of the vestibular nerve projections from the posterior canal on the other side. |
|
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
Specific pairs of ocular muscles are stimulated by each canal pair. (Review the table in Ch. 7 of the cranial nerve projections.) These muscles have insertion planes that parallel the canals that stimulate them. The crossed innervation of the SR and SO allows the vestibular system to compensate for vertical head rotation with vertical movements of both eyes by stimulating only one side of the brainstem. Downward head rotation stimulates the anterior canal and innervates the contralateral III causing innervation of the ipsilateral SR and contralateral IO. Upward head rotation stimulates the posterior canal and also innervates the contalateral trochlear IV and contralateral III causing innervation of the ipsilateral SO and contralateral IR. The horizontal canal projects to the ipsilateral vestibular nucleus and then to the contralateral abducens which innervates the contralateral LR. The abducens also sends a crossed interneuron back to innervate the ipsilateral III and MR. |
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
Table 13.1 Yoked muscles innervated by vestibular canals |
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
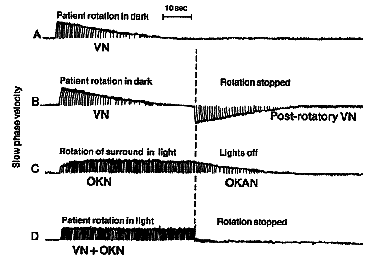
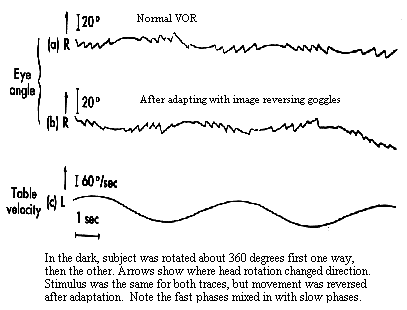
The VOR responds too quickly to utilize visual feedback to check its accuracy. Nonetheless it is extremely accurate. This precision results from long term calibration or adaptation to conditions that change the relation between head rotation and resulting velocity of the retinal image. Changes are produced with spectacle lenses or anomalies affecting the mechanical transducer in the canals. Adaptation is accomplished during visual stimulation when errors or residual retinal slip can be detected. These visual errors are sent to the cerebellum along with sensed measures of eye movement called efference copy. Taken together retinal image motion and sensed eye motion give us a sense of the motion of a target in space. We need this information if we want to follow a moving target with our eyes. Initially the eye is stationary and the target is moving. The eye senses the motion from the retinal image and starts following. This following stabilizes the retinal image at the fovea. What keeps the eye following the target once this stabilization has been established and there is no more retinal slip? The stimulus is perceived target motion that comes from our sense of eye movement. Adaptation of the VOR occurs by changing our sense of eye movement. If we want a faster VOR we exaggerate our sense of eye movement velocity and this produces a higher stimulus velocity to drive the following reflex. This modification occurs in the vestibular nuclei and its interactions with the flocculus. The VOR is so plastic that it can even respond to magnifying lenses and reversing prisms. Those of you who wear glasses should look across the room and shake your head horizontally. When you do this the world will appear stable. Now do the same thing with your glasses off. If you are a myope you are used to looking through minifying lenses and your VOR is reduced. Without your glasses the world appears to move in the opposite direction as your head. If you are a hyperope, your plus lenses have magnified the retinal image and increased the amplitude of your VOR. Without your glasses the world appears to move in the same direction as your head. In an extreme experiment, Stratton, the founder of Berkeley's Psychology Department, wore 180 deg inverting prisms in the form of an astronomer's telescope for two weeks. After that period, his VOR was reversed so that when his head rotated to the right, his eyes did as well. It is a very remarkable system. |
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
Opthalmostatic reflex |
||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||
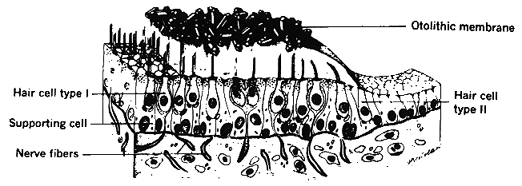
Fig 13.10 Otholith organ (located in floor of utricle) |
||||||||||||||||||||||||||||||||||||||||
In animals such as the rabbit with laterally placed eyes, the roll of the head stimulates the urticle and causes upward rotation of one eye and downward rotation of the other. These are called skew movements and they are accompanied by ocular torsion. The same motion of the globe in man is called the Ocular Counter Roll (OCR) reflex. Upward motion of one eye in man is also referred to as a skew movement. Stimulation of one utricle produces skew like movements of the ipsilateral eye similar to those seen in See-Saw nystagmus where the ipsilateral eye intorts as it moves up and the contralateral eye extorts as it moves down. There is also the ocular tilt response (OTR) in which the same skew-torsion response results from stimulation of the contralateral nucleus of Cajal. A different pattern of torsion and skew is seen in the occlusion hypertropia first observed by Bielchowsky. Occlusion of either eye causes an elevation and extorsion of the covered eye. However, more commonly it is referred to as a hyper deviation. These are often accompanied by torsion of the eyes and are likely to be involved in the double occlusion hyperphoria or dissociated vertical deviation (DVD) also known as the Bielchowsky darkening sign. The vestibular organs are mainly used for a sense of balance and equilibrium, however we also use vision for this purpose. To demonstrate this try to stand on one foot with your eyes open and then closed. It is very difficult to do this without visual stimulation. The visual sense is also noted in the stereotyped imbalance of a person standing on a cliff or edge of a building. They lack the visual sense of the ground plane to give them posture stability. The strength of the visual input can override signals from the vestibular system in situations like the ìmystery houseî that is really built on a hill at an angle from true (gravitational) vertical. The conflict is resolved by seeing water flow uphill and people stand at oblique angles. This is a rather subjective phenomenon. Astronauts in space are perfectly happy to float upside down with respect to the rest of the instruments in the cabin, however if they see another astronaut floating in an inverted orientation with respect to themselves they may throw up. Both visual and vestibular signals are fed into the floccular or vestibular cerebellum for sensing orientation and guiding posture control. |
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
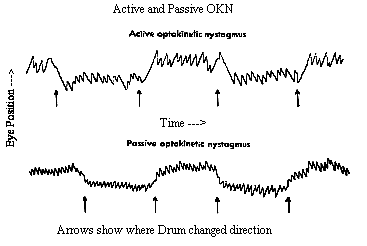
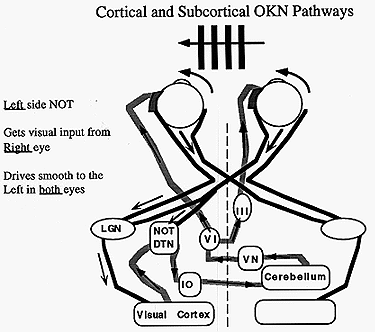
The OKR is very similar to pursuit tracking and is often confused. For example it is possible to foveally track a single feature in the moving field or let the eye reflexively take over the OKN response. These two modes of smooth response can be distinguished by their fast phase frequency and by the purpose of the fast phase. Repetitive pursuit tracking can only occur 1-1.2 times/second whereas reflex OKN has a reset rate of 3 Hz. In addition, when pursuing, the slow phase response follows the target as it moves into the periphery and then the saccade resets the eye into primary gaze. In contrast, reflex OKN makes a saccade away from primary gaze into the direction the field is emerging from and then the eyes move smoothly toward primary gaze. In part, this is an alert response where the eyes move toward a novel moving target in the periphery and follow it in toward primary gaze. The names given to these two modes of following are look or active OKN for the pursuit response, and stare or passive OKN for the reflex response. These names come from instructions to subjects to elicit each response. Subjects are told either to look at a single feature and follow it, or to stare at the whole field and keep it visible without trying to follow it intentionally. All parts of the retina are sensitive to stare OKN but the central foveal area is most sensitive. OKN is also responsive to large fields as opposed to small ones less than 20 degrees. The upper stimulus velocity we can follow increases with field size up to 50 degrees. The maximum velocity rarely exceeds 50 deg/sec and usually is close to the stimulus velocity below 30 deg/sec. If OKN continues for about 1 minute and then the subject sits in darkness, the OKN continues for about 30 seconds as optokinetic afternystagmus. Similar effects occur with the VOR once the endolymph has settled down after rotation stops. (See Fig 13.7) These aftereffects are a means for the VOR and OKN to continue responding even to constant velocity head rotation when the head stops accelerating while rotating. The aftereffect is sometimes called velocity storage and its site of action is the vestibular nuclei. Afternystagmus has two stages. The first stage is one in which the OKN continues in the same direction as during light stimulation. The second phase is one in which the nystagmus reverses direction. This second stage reveals a habituation mechanism that is intended to slow down or cancel OKN during long periods of optokinetic stimulation. Its function might be to suppress the reflex to allow the eyes to view moving targets that are not fixed with respect to the background. The visual stimulus for posture control is derived from optic flow of the retinal image. The retina contains ganglion cells that respond exclusively to motion in certain directions or orientations. This information passes along the optic nerve, decussates at the chiasm and goes to the cortex via the geniculate body (LGN) or to the midbrain via the accessory optic tract. This tract has several nuclei in the pretectal area. One pair of these nuclei, the nucleus of the optic tract (NOT), is tuned to horizontal target motion. The lateral terminal nuclei (LTN) are tuned for vertical OKN. Neurons in these nuclei have large receptive fields and respond to large textured stimuli moving in specific directions. Stimulation of the right NOT with rightward motion causes following movements of the eyes to the right or ipsilateral side, and similarly stimulation of the left NOT with leftward motion causes leftward following movements. (See Fig 13.12 below) Each NOT projects signals via the inferior olive to the vestibular nuclei and possibly to the flocculus via the climbing fibers of the cerebellum (monkey but not man). (Reference: page 162 Adler). |
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
The organization of the afferent projections to the NOTs can be used to interpret developmental changes in OKN during infancy. Each NOT receives two sources of input. One is a direct subcortical projection from the nasal retina of the contralateral eye and the other is a cortical-tectal projection originating from the temporal retina of the ipsilateral eye. In addition there are crossed projections conveying nasalward target motion that go to the cortex via the LGN and then descend to the NOT with the ipsilateral uncrossed projection from the contralateral eye. (Refer to Fig 13.12 above) The cortical region that organizes motion signals is the MST, medial superior temporal lobe. This region is important for generating motion signals for both pursuits and OKN. There are binocular cells in the NOT that merge the crossed subcortical-retinal projection with uncrossed cortical projections of the other eye. (A schematic of these pathways is shown in the middle of Fig 5-84 in Adler's physiology textbook.) Until the age of 3 months, the subcortical projections predominate because the cortical projection is not yet developed. Thus the only way we can stimulate OKN in an infant is from the subcortical crossed input. The result is during monocular stimulation we can stimulate a nasalward slow phase of OKN (both eyes move smoothly toward the covered eye) but not a temporalward slow phase because the cortical projections to the tectum that convey temporal motion information have not yet developed. Binocular stimulation causes OKN to respond to either left or right motion because the motion is nasalward for one or the other eye and this results in a subcortical stimulation of the NOT. After 3 months the infantís cortical projections predominate and horizontal OKN responds to both temporalward and nasalward image motion. |
||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||
An interesting problem occurs in infantile strabismus. These patients fail to develop normal binocular vision because of the strabismus. Their lack of cortical binocularity has a secondary effect which is to disrupt the cortical tectal pathways that convey temporalward motion information originating from the ipsilateral eye to the tectum. In strabismus, this results from less representation of the ipsilateral than contralateral eye in each cortical hemisphere. The loss of ocular dominance of the ipsilateral eye is caused by developmental competition for cortical dominance by the two eyes and suppression of the ipsilateral eye in strabismus. Thus patients with infantile strabismus have the same asymmetric OKN pattern as in immature infants. As mentioned above, these patients also have latent nystagmus which appears to be a manifestation of the asymmetric OKN. During monocular fixation, the stimulated retina increases the activity of neurons in the contralateral NOT via subcortical crossed projections, but it has no way to innervate the ipsilateral NOT. The result is the eyes position is pulled to the side of the stimulated NOT which is the side of the covered eye. The fixation error is corrected with a saccade and repeated sequence becomes latent or occlusion nystagmus. Review Questions:
|
||||||||||||||||||||||||||||||||||||||||
[Previous Chapter] [Next Chapter] |
||||||||||||||||||||||||||||||||||||||||