[Table of Contents] [Home] [Glossary] [Video] [Help]
THE CEREBELLUM:
OCULOMOTOR REPAIR SHOP
Key Words: cerebellum, flocculus, nodulus, vermis,
Purkinje cell
Outline
VIII. Cerebellum- The oculomotor repair shop
- Introduction
- Functions
Breaking/ damping movements
Predicts position
Generates reflex templates
Compensates for errors
- Gross Anatomy
Vestibulo-cerebellum (flocculus/ nodulus) -- smooth eye movements OKN, Pursuits; step component of saccades
Vermis -- controls the pulse component of saccades
- Cellular Architecture
Input from afferent sources via climbing fibers and mossy fibers
Output from Pukinje cells which project to various supranuclear relay stations
Interconnection between in and output by granular cells (T-junction)
Output is exclusively inhibitory
- Short-term Adaptation
Compares intended eye movement (afferent and efferent correlates) with sensed eye movement (reafference)
Adaptation at granular cell input to Purkinje cell output layer
- (Outline Continued in Chapter 11)
Introduction
The cerebellum is known as the posterior brain. It
is Latin for little brain and it is vital for the control of
very rapid movements such as running, talking and fast eye movements
like saccades. It is a comparator. It monitors and
makes correct adjustments in the motor activities elicited by
other parts of the brain. It receives continuous updates
about vestibular, proprioception, vision, somatosensory, auditory,
head and eye movements to determine instantaneously the status
of position and motion of body limbs and eye position. The cerebellum
compares the actual physical status of a body part as indicated
by sensory information to intended responses of the cortex. Discrepancies
are then corrected with instantaneous feedback and adaptive long
term changes. Control by the cerebellum modulates activity
in the brainstem using inhibition by Purkinje cells. There are
deep nuclei beneath the cerebellar lobes that serve as relay
stations for the output of Purkinje cells.
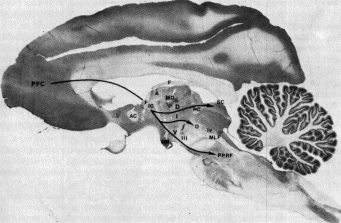
|
Fig 10.1 Monkey brain. Note the posterior position of cerebellum (leaf-like structure in cross section) (Projections from prefrontal cortex to motor structures in the midbrain are labeled in this illustration.) |
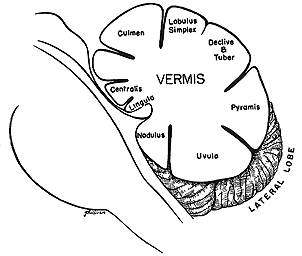
|
Fig 10.2 Cerebellum |
|
1) Breaking or damping: Regarding fast movements, the cerebellum can exert or initiate breaking impulses to slow down or dampen a movement. Ordinarily, the motor cortex transmits far more impulses than are needed to perform a movement and the cerebellum inhibits the motor cortex at the appropriate time. The cerebellum calculates the length of time that is required to reach the point of intent (time to contact). It breaks by stimulation of the antagonist and inhibition of the agonist. Stimulation of the antagonist is entirely automatic. This dampens the eye at the end of the Saccade. If there were no damping saccadic overshoots and oscillations would occur (ocular flutter).
2) Prediction: The cerebellum helps predict future positions of a moving eye and stops the eye when it reaches its final position in less time than 1 reaction time. Without prediction, the eye would overshoot its mark and dysmetria would occur. The cerebellum is also used in locomotion to predict time of contact and when approaching a wall or stop sign in a car. If you remove the cerebellum in a monkey it runs into walls.
3) Reflex templates or engrams: The cerebellum generates reflex following movements to stabilize vision during locomotion. The repetitive wave form in jerk nystagmus is generated by the cerebellum by comparing ongoing eye movements with the stored wave form and differences are sensed and used to generate nystagmus to match the template.
4) Adaptive long-term compensation for errors in motor activity: The cerebellum can adjust gain or sensitivity of neurons in the vestibular nuclei to adjust the amplitude of movements of the eyes during OKN and the VOR. Both the amplitude and phase of responses can be adjusted. For example if you wear magnifying plus spectacle lenses, the images are magnified and the eyes must rotate more than the head to stabilize the retinal image. This is accomplished with the aid of the cerebellum. Also, if you invert vision with an astronomer's telescope (like Stratton did at Berkeley at the turn of the century), the visual field appears to rotate with the head rather than in the opposite direction as it normally does. After wearing this apparatus for one week, the VOR is reversed, i.e. when you rotate your head to the right the eyes also rotate to the right, not to the left as they normally do. This constitutes a phase reversal of this reflex motor response.
Gross Anatomy
The cerebellum is divided into three functional divisions: the vestibulocerebellum (consisting of the flocculus and nodulus), the spinocerebellum (consisting of the vermis and the intermediate part of the hemispheres), and the cerebrocerebellum (consisting of the lateral part of the hemispheres).
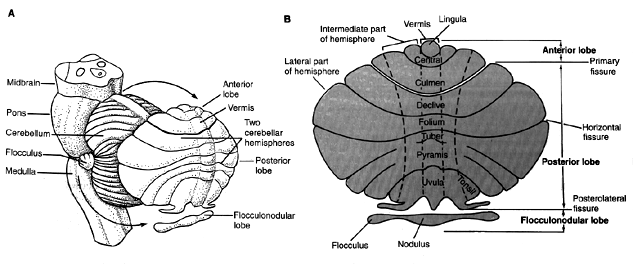
Note actual locations of structures such as the vestibulocerebellum (flocculus and nodulus) in ìAî
Phylogenetically, the oldest region is the vestibulocerebellum, which is responsible for maintaining equilibrium. It is made up of the flocculus and nodulus in the rostral, ventral region of the cerebellum. This region gets input from and passes information to the vestibular nuclei. It gets head position input from the vestibular system ( the semicircular canals which signal head acceleration, the otoliths that signal head orientation with respect to gravity) and neck proprioception. It also gets visual input from the LGN and the accessory optic tract via the inferior olive, superior colliculi, striate cortex via the lateral dorsal pontine nuclei and eye position information from the prepositus hypoglossi. Purkinje cells fire in relation to gaze velocity or velocity of the eye in space during pursuit of a moving target with the head still or during combined eye-head rotation. It governs body equilibrium during stance and gate, smooth eye movements in OKN and pursuits, adaptation of the step component of saccades, and adaptation of the gain and phase of the VOR to spectacle lenses and near viewing distances. Anomalies resulting from lesions in the flocculus are nystagmus, cogwheel pursuits, low velocity (gain) OKN, equilibrium disorders, step-pulse mismatch resulting in glassades, inability to hold gaze away from primary position, and inability to adapt the gain of the VOR or to suppress the VOR during pursuits. Lesions in the nodulus affect vertical OKN and cause periodic alternating nystagmus. (p.41 in Zee).
Vermis
The vermis lies along the midsagittal region of the cerebellum. It receives peripheral information from the spinal cord, auditory, visual vestibular and somatosensory and motor cortices for peripheral body representation. A homonculus of the whole human body is represented in the vermis. In terms of eye movements it receives proprioceptive information from the eye movements. The vermis adjusts lower brainstem mechanisms which determie the size of a saccade. Lesions cause saccade dysmetria; this indicates that the vermis keeps a central record of eye position which is updated by both proprioceptive and visual information. The central record may determine how far the eyes would be allowed to deviate from primary position before they were reset by a saccade. Lobes VI and VII are responsible for regulating the pulse component of saccades, breaking the saccade at the end of the movement and for integrating head and eye movements and retinal image motion (slip) during pursuit. The vermis is supposed to guide pursuits to changing velocity whereas the flocculus maintains pursuit during steady tracking. Anomalies resulting from lesions in this region include saccade dysmetria including both hypermetric and hypometric saccades, impaired pursuits while changing velocity, tremor of limbs and loss of muscle tone.
Cellular Structure and Organization
The cerebellum may be called the ìlittle brainî but it contains more cells than the remaining brain. In particular, one small class of cells called granular cells number more than 100 billion. There are as many granular cells as there are stars in the sky. The surface of the cerebellum is extremely orderly and uniform. The same types of cells are found everywhere, they only perform different tasks. The surface grey matter can be divided into 3 layers, as illustrated in Figure 10.4, below:
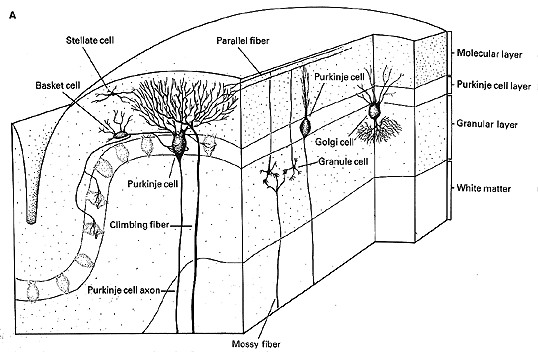
|
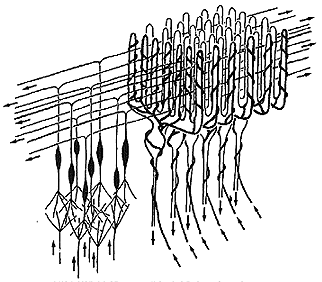
Fig 10.5 Cell and fiber types shown here are: (Refer to Fig 10.6 below to identify cell types.) |
|
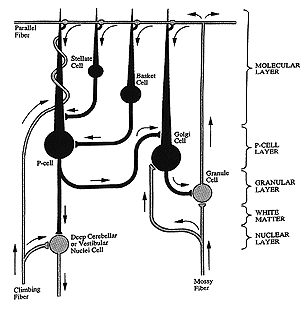
Fig 10.6
|
There are two classes of excitatory inputs to the cerebellum. One comes exclusively from the inferior olive and relays visual optic flow from the accessory optic tract. This input is called the climbing fiber and it literally climbs around an individual Purkinje cells like ivy on a telephone pole. (See figures 10.5 and 10.6 above) One climbing fiber contacts 10 Purkinje cells and each Purkinje cell only receives input from one climbing fiber. Thus each climbing fiber has its own 10 Purkinje cells. The other input is the mossy fiber that originates from all other sources including the vestibular nuclei. One mossy fiber contacts 1000 Purkinje cells.
Adaptation
The adaptive nature of the cerebellum arises from
the connections made between excitatory input fibers and output
inhibitory Purkinje cells. The cerebellum compares intended
eye movement using efference copy with the actual movement of
the eye, sensed with visual feedback, and makes adjustments in
the efferent flow of innervation originating in supranuclear
regions such as the vestibular nuclei. Apparently the cerebellum
doesn't use stretch receptor inputs for eye movement control
but rather an intended innervation that is calibrated using stretch
receptor information over a long term process of learning (1
week or so). It acts like a regulator or valve for controlling
the strength of innervation. The strength of innervation
is determined with inhibition by Purkinje cells that varies with
the number of connections to the Purkinje cell provided by the
granular cells. The granular cell connections are modified
to include more or fewer synapses to change the amount of inhibition
applied to supranuclear sites.
Review Questions:
- Describe the cellular layers of the cerebellum and their functions.
- What input to the cerebellum is primarily visual?
- What is the primary influence of the Purkinje cells.
- What are the roles of the vermis and vestibulo-cerebellum?
[Table of Contents] [Home] [Glossary] [Video] [Help]