Home Page
James Maxwell
 |
PublicationsSelected Abstracts |
Research Interests:
 Vergence eye movements Saccade-vergence interactions Noncomitant vertical phoria adaptation
Vergence eye movements Saccade-vergence interactions Noncomitant vertical phoria adaptation
Otolith-dependent adaptation of vertical skew
Neurophysiology of binocular eye movements
 |
Contact information:James S. MaxwellSchool of optometry 360 minor hall University of california Berkeley, ca 94720-2020 Phone: (510) 642-7710 Fax: (510) 643-5109 E-mail: jsm@garnet.berkeley.edu |
PUBLICATIONS:
- Maxwell, J.S. and Schor, C.M. Head-position-dependent adaptation of nonconcomitant vertical skew. 37 (4), 441-446, 1997.
- Maxwell, J.S. and Schor, C.M. Adaptation of vertical eye alignment in relation to head tilt. Vision Research, 36 (8), 1195-1205, 1996.
- McCandless, J.W., Schor, C.M., Maxwell, J.S. A cross-coupling model of vertical vergence adaptation. IEEE Transactions on Biomedical Engineering. 43 (1), 24-34, 1996.
- King, W.M., Zhou, W., Tomlinson, R.D., McConville, K.M.V., Page, W.K., Paige, G.D., Maxwell, J.S. Eye-position signals in the abducens and oculomotor nuclei of monkeys during ocular convergence. J. Vestibular Research. 4 (5), 401-408, 1994.
- King, W.M., Zhou, W., Tomlinson, R.D., McConville, K.M.V., Page, G.D., Maxwell, J.S. Eye-position signals in the abducens nucleus of monkeys during ocular convergence. In, Contemporary Ocular Motor and Vestibular Research: A Tribute to David A. Robinson. Fuchs, A., Brandt, T., Buettner, U., Zee, D., (eds.) Thieme, Stuttgart, 1994.
- Schor, C.M., Maxwell, J.S. and Stevenson, S.B. Vertical conjugacy and the theoretical isovergence surfaces. In, Contemporary Ocular Motor and Vestibular Research: A Tribute to David A. Robinson. Fuchs, A., Brandt, T., Buettner, U., Zee, D., (eds.) Thieme, Stuttgart, 1994.
- Schor, C.M., Maxwell, J. S., Stevenson, S.B. Isovergence surfaces: The conjugacy of vertical eye movements in tertiary positions of gaze. Ophthalmologic and Physiological Optics. 14, 279-286, 1994.
- Gleason, Schor, C., Lunn, R., Maxwell, J., Directionally selective short-term nonconjugate adaptation of vertical pursuits. Vision Research. 33, 65-71, 1993.
- Maxwell, J.S. and Schor, C.M. Mechanisms of vertical phoria adaptation revealed by time course and two dimensional spatiotopic maps. Vision Research. 34, 241-251, 1993.
- Schor C., Gleason, G., Maxwell, J., Lunn, R., Spatial aspects of vertical phoria adaptation. Vision Research. 33, 73-84, 1993.
- Maxwell, J.S. and King, W.M., The dynamics and efficacy of saccade-facilitated vergence eye movements in monkeys. J. Neurophysiology. 68, 1248-1260, 1992.
MANUSCRIPTS:
Maxwell, J.S. The interaction of saccades and vergence eye movements and the effect of vergence angle on the discharge rate of abducens neurons in the monkey. Ph.D. Thesis, University of Rochester, Rochester, NY, 1991.Abstracts for selected publications
Mechanisms of Vertical Phoria Adaptation Revealed by Time-Course and Two-Dimensional Spatiotopic Maps, James S. Maxwell and Clifton M. Schor, 1994.
The spatial spread of short-term phoria adaptation was measured in response to either a single vertical disparity presented at a single eye position, or, vertical disparities of opposite sign presented at two different locations along either the primary vertical or horizontal meridians or along an oblique axis. The spread of adaptation to eye positions not specifically adapted was assessed by measuring phoria across a two-dimensional surface. The change in phoria was uniform across the field in response to a single disparity. With two disparities, adaptation conformed to the stimulus demand in the direction in which the disparity varied but was uniform in the orthogonal direction. The time course of the adaptation indicated the presence of two mechanisms, a global one which shifted the phoria uniformly across the field and a local one which selectively adjusted the phoria to the position-dependent demands of the disparity.
Adaptation of Vertical Eye Alignment in Relation to Head Tilt, James S. Maxwell and Clifton M. Schor, I996
Binocular visual feedback is used to continually calibrate binocular eye alignment so that the retinal images of the two eyes remain in correspondence. Past experiments have shown that vertical eye alignment (measured as vertical phoria) can be altered by training to disparities that vary as a function of orbital eye position. The present experiments demonstrate that vertical eye alignment can also be trained to differ with head position when eye position (with respect to the orbit) is held constant. Changes in head position were about either an earth-vertical or earth-horizontal axis to distinguish otolith-ocular related adaptation from cervical-ocular related adaptation. Changes in head position were implemented by either by rotating the whole body (WB) or by rotating the head with the body stationary (HO). Following training, adaptation of eye alignment was observed in all cases of rotation about an earth-horizontal axis and for HO pitch rotations about an earth-vertical axis. The results illustrate the ability of the oculomotor system to compensate for imbalances in otolith-ocular pathways.
Head-position-dependent adaptation of nonconcomitant vertical skew, James S. Maxwell and Clifton M. Schor, 1997
Vertical phoria can be trained to vary with either head position or orbital eye position. The present experiments show that subjects can simultaneously adapt their eye-position-specific (noncomitant) vertical phorias in different directions at different head positions. Eye-position-dependent and head-position-dependent adaptive pathways, therefore, are not independent. Rather, the adaptation of vertical skew takes into account both eye and head position. In additional experiments, the magnitude of the noncomitant adaptive response was shown to be related to otolith output, increasing with head tilt ipsilateral to the tilt position at which training was received and decreasing in the contralateral direction.Dynamics and Efficacy of Saccade-Facilitated Vergence Eye Movements in Monkeys,J.S. Maxwell and W.M. King, 1992
Macaque monkeys were trained to alternately fixate targets requiring different combinations of horizontal version and vergence. Eye movements were recorded binocularly with search coils. The peak vergence velocity for combined vergence-saccade eye movements reached higher velocities than for vergence movements alone and vergence speed increased in direct proportion to saccade amplitude. We determined, however, that most of the increase in speed was due to the addition of transient vergence movements that occur for all saccades, even those between targets at the same distance where no vergence is required. Even so, vergence speeds during combined vergence-saccades exceeded the speeds predicted by the linear addition of saccade-related vergence transients and pure vergence movements. The increased vergence speeds were behaviorally relevant in that vergence duration was decreased. A distinction was made between the intrasaccadic vergence that occurred during saccades and the intrasaccadic vergence that occurred because of saccades by calculating the amount of vergence that would have occurred during the duration of the saccade (based on the average speed of pure vergence movements of comparable size and assuming linear addition) and subtracting that from the actual vergence obtained. The difference was assumed to be the portion of vergence contributed by a saccade-related mechanism and not by the slow vergence controller.
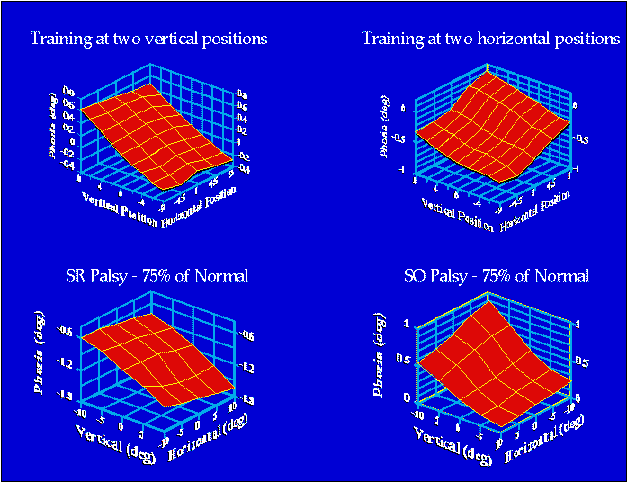 The upper plots show the average phoria for four subjects
following training in the Purkinje eye
tracker. The left-hand figure shows the result of training with a right
hyperdisparity (right eye's image higher than the left's) in the upper field
and a left hyperdisparity in the lower field for forty minutes. The right-hand
plot shows the result of training with a right hyperdisparity in the right
field and a left hyperdisparity in the left field. Phoria was measured by
having one eye occluded while the other viewed an array of targets. The
lower plots show eye alignment as predicted by a model of orbital mechanics
(Orbit TM given a
75% palsy of the superior rectus (left figure) or superior oblique (right
figure). These simulations demonstrate that one purpose of an eye-position-dependent
eye-aligment adaptive mechanism would be to compensate for misalignments
produced by palsies of the extraocular muscles.
The upper plots show the average phoria for four subjects
following training in the Purkinje eye
tracker. The left-hand figure shows the result of training with a right
hyperdisparity (right eye's image higher than the left's) in the upper field
and a left hyperdisparity in the lower field for forty minutes. The right-hand
plot shows the result of training with a right hyperdisparity in the right
field and a left hyperdisparity in the left field. Phoria was measured by
having one eye occluded while the other viewed an array of targets. The
lower plots show eye alignment as predicted by a model of orbital mechanics
(Orbit TM given a
75% palsy of the superior rectus (left figure) or superior oblique (right
figure). These simulations demonstrate that one purpose of an eye-position-dependent
eye-aligment adaptive mechanism would be to compensate for misalignments
produced by palsies of the extraocular muscles.
 Cliff is demonstrating the three axes of rotation that
were used in this experiment (Please note that his head was not actually
removed from his body). The head was stationary during training and testing.Why
is Cliff wearing the stylish glasses?
Cliff is demonstrating the three axes of rotation that
were used in this experiment (Please note that his head was not actually
removed from his body). The head was stationary during training and testing.Why
is Cliff wearing the stylish glasses?
 Subjects (none of them were actually bald headed by
the way) trained their vertical phorias by looking through an afocal magnifier
with one eye. The magnifier was in front of one eye with the head rolled
to the right and in front of the other eye when the head was rolled to the
left. The magnification makes that eye's image appear higher than the other
when viewing a target in the upper visual field and lower than the other
when looking at a target in the lower field. After one hour of training
the subjects' vertical phorias changed in a way that would eliminate the
double vision initially introduced by the lenses.
Subjects (none of them were actually bald headed by
the way) trained their vertical phorias by looking through an afocal magnifier
with one eye. The magnifier was in front of one eye with the head rolled
to the right and in front of the other eye when the head was rolled to the
left. The magnification makes that eye's image appear higher than the other
when viewing a target in the upper visual field and lower than the other
when looking at a target in the lower field. After one hour of training
the subjects' vertical phorias changed in a way that would eliminate the
double vision initially introduced by the lenses.